
The Final Nail
A Response to Criticism of the Phase 1 Physical-Evidence Falsification of Modern Virology

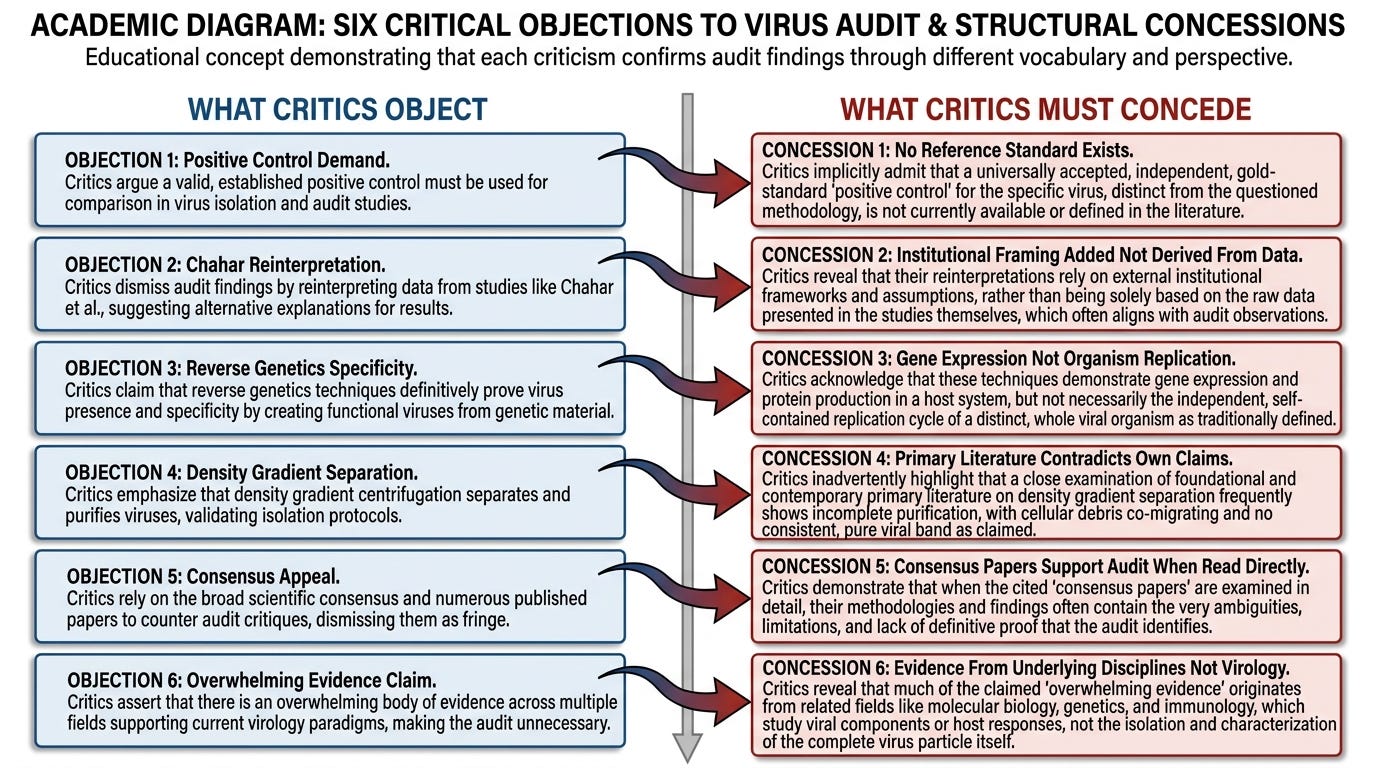
What the Critics Have to Concede
Companion framework: The Open Reality Bible v2.9.49 (§§5.6.U, 5.6.P, 5.6.R, 5.6.S, 5.13.E, 5.13.I, 5.14, 2.7, 2.13)
The Phase 1 paper — A Complete Physical-Evidence Falsification of Modern Virology — made one argument, sustained from 1892 to the present-day laboratory. This response addresses the criticisms that argument predictably attracts. Not the dismissals. The actual technical objections that a careful institutional defender would raise, and what those objections require the defender to concede in order to make them.
Each objection, examined honestly, requires the defender to accept the foundational audit finding. The objections do not save the institutional model. They confirm its structural problem in different vocabulary.
Objection One: "Lanka did not run a proper positive control."
The institutional criticism of the April 2021 Lanka experiment is methodological. Lanka did not include a parallel condition in which cells were exposed to authenticated SARS-CoV-2 virus stock and showed the expected cytopathic effect, against which his control conditions could be measured. The absence of the positive control, the argument runs, means the experiment cannot conclude that CPE is non-specific to viral presence.
This objection is unmeetable on its own terms. A proper positive control, by the standards of experimental design, requires a known-positive reference standard. The known-positive reference standard for "virus presence" must be a purified, characterized, discrete viral particle in defined concentration. No such reference standard exists. Per the institutional framework's own admissions across the relevant primary literature, SARS-CoV-2 has never been physically isolated as a discrete particle, purified to homogeneity, characterized independently of cell-culture amplification, and made available as a reference standard for positive-control experiments.
What the institutional framework calls a positive control is, structurally, cells exposed to stressed-cell supernatant labeled as containing virus. The "virus stock" is itself the product of the cell-culture protocol whose specificity is the question under audit. Using stressed-cell supernatant as the positive control to validate that cell-culture stress produces CPE is not a control. It is the experimental condition tested against itself.
The critic demanding that Lanka use a "proper positive control" is demanding that he use a reference standard the institutional framework cannot produce. The demand is unmeetable not because Lanka failed to meet it but because the reference standard does not exist. To press the objection, the critic must either provide the missing reference standard — which would resolve the entire identification question independently — or concede that the institutional positive controls are doing the same thing Lanka's experiment did, with the labels left on.
What the critic must concede: that "positive control" in virology, as currently practiced, does not contain a discrete characterized viral entity, and that the institutional protocol cannot meet the standard the critic is demanding of the audit.
Objection Two: "You are misreading Chahar et al."
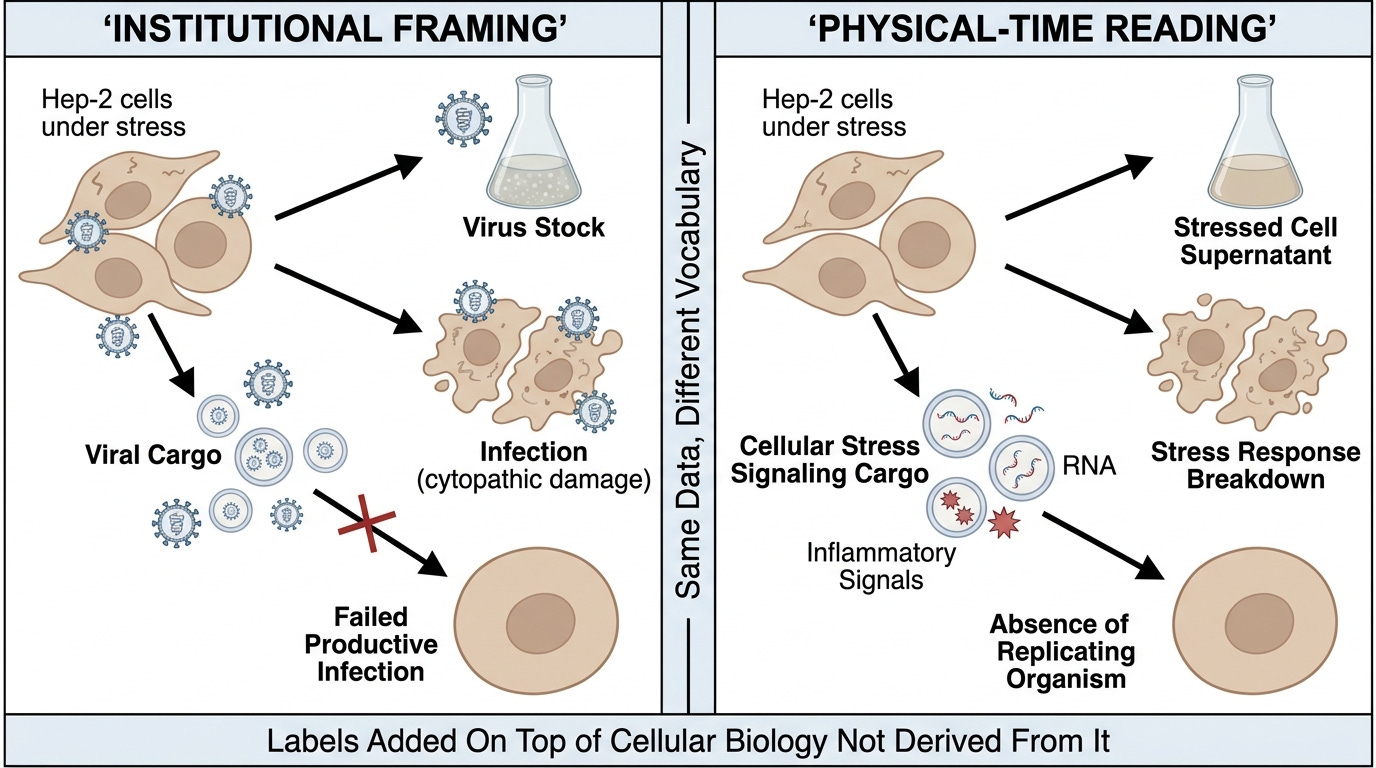
The audit reframes the 2018 Chahar paper. Where the authors describe their finding as exosomes carrying viral cargo that fail to establish productive infection, the audit reads the same protocol as stressed-cell exosome biology accurately documented in molecular and cellular terms while framed within an institutional vocabulary that preserves the discrete-organism model. The critic objects that this is reinterpretation against the authors' own conclusions.
The objection assumes the authors' conclusions and the experimental data are the same thing. They are not. The authors performed a sequence of cellular and biochemical operations and reported results. Those results — fragmented RNA in exosomes, partial proteins, inflammatory signaling cargo, absence of productive infection in receiving cells — are the data. The framing of the data within the institutional model is the interpretation. The data is fixed. The interpretation is what the audit is testing.
The audit-honest reading of the Chahar protocol, stripped of the institutional framing, is the following. Stressed cells (Hep-2 cells in standard culture stress conditions) produced supernatant. The supernatant caused cytopathic damage in fresh stressed Hep-2 cells. The damage count was operationally defined as a "virion titer." The operationally-defined "virion stock" was added to A549 cells under matched stress conditions. The A549 cells responded with stress and breakdown — the expected response to additional stress input. Supernatant from the stressed A549 cells contained exosomes carrying fragmented cellular material and inflammatory signals. These exosomes, when added to non-stressed cells, propagated inflammatory signaling but did not cause cytopathic breakdown — because non-stressed cells receiving stress signals are not the same as stressed cells receiving more stress.
Every step is real cellular biology, documented at the physical-time layer by the paper's own methods. What changes between the institutional reading and the audit reading is what the steps are called. The institutional reading calls the supernatant "virus stock," the cellular damage "infection," the inflammatory cargo "viral cargo," and the final non-transmission "failed productive infection." The audit reading calls the supernatant stressed-cell supernatant, the cellular damage stress-response breakdown, the inflammatory cargo cellular stress signaling, and the final non-transmission an absence of replicating organisms to fail.
The audit is not reinterpreting the data. It is reading the data without the relabeling. The critic objecting to the audit reading must defend the relabeling as adding something beyond what the cellular biology already documents. The defense is unavailable because the labels were added on top of the cellular biology, not derived from it.
What the critic must concede: that the institutional framing of the Chahar paper does not derive from the cellular and molecular data the paper actually contains, but is applied to that data from outside.
Objection Three: "Reverse genetics with sequence-specific mutations proves the virus is real."
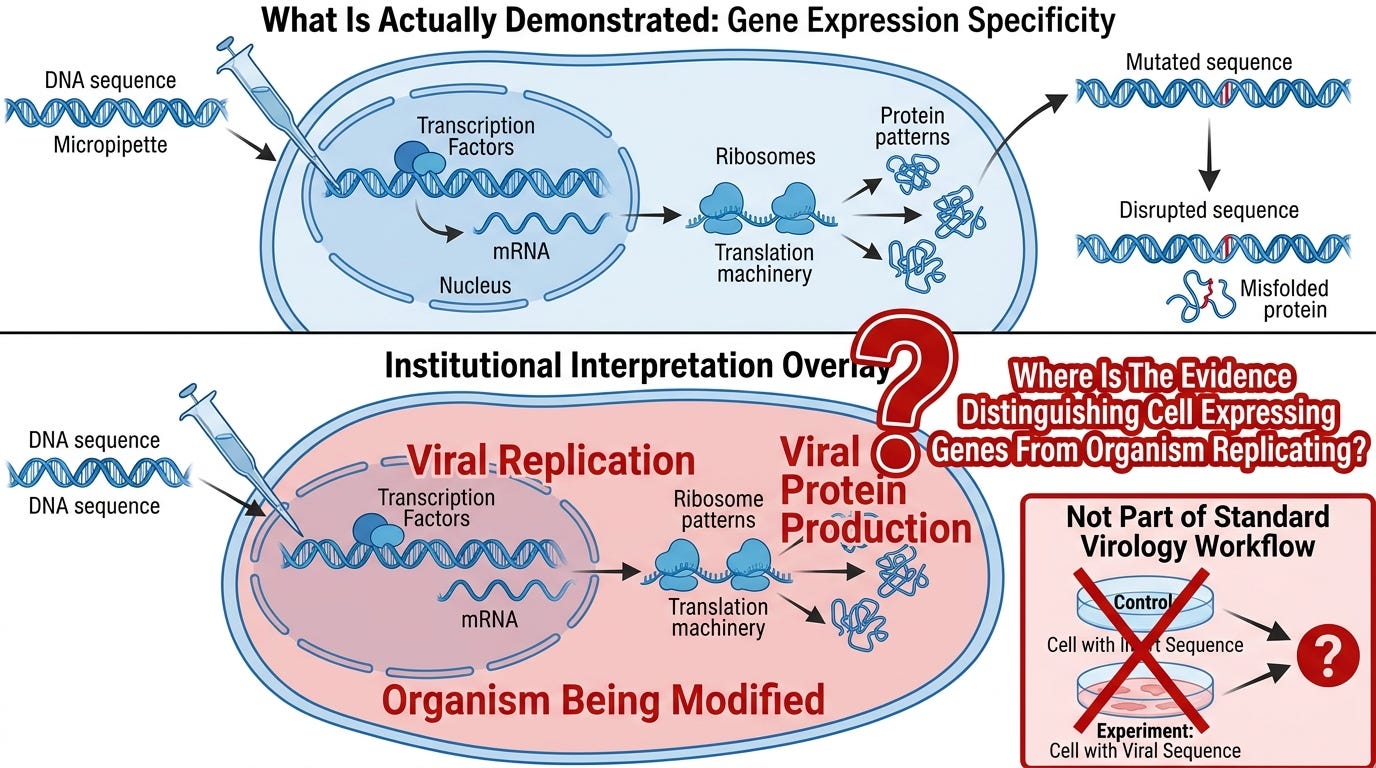
The critic's strongest experimental claim is that reverse-genetics workflows with sequence-specific genetic modifications produce sequence-specific phenotypic outcomes. A stop codon introduced mid-genome reliably eliminates the downstream effect. A polymerase knockout reliably eliminates replication signals. A receptor-binding-domain mutation reliably alters receptor binding. The specificity, the critic argues, demonstrates that a specific organism is being manipulated.
This objection commits a category error so structural that it explains the entire field. Sequence-specific phenotypic outcomes in cell-culture experiments are not evidence of organism replication. They are evidence of gene expression specificity. The cell reads the genetic input, transcribes it, translates it, and produces the encoded products in sequence-specific patterns. Mutations that disrupt coding sequences produce predictable, reproducible disruptions in protein output. This is molecular biology working exactly as molecular biology characterizes it. Genes encode proteins. Sequence changes alter the encoded proteins. The cellular machinery responds in sequence-specific ways to sequence-specific inputs.
The institutional framing labels this "viral replication" and "viral protein production" and treats sequence-specific responses as evidence that a specific organism is being modified. The audit-honest framing is that sequence-specific cellular responses to sequence-specific genetic inputs are exactly what molecular biology says cells do. The specificity is real and reproducible. What the specificity establishes is gene expression specificity — not organism replication.
The discipline of molecular biology developed the experimental tools that reverse-genetics workflows use. Those tools were developed to characterize gene expression, regulation, and protein production. They do that work rigorously. Virology imports the tools and the data they produce and adds an entity-replication interpretation that the tools were not designed to test. The reverse-genetics chain documents what cellular machinery does with introduced genetic inputs. It does not document organism replication, because the experimental design does not distinguish "the cell expressing the encoded products" from "an organism reproducing itself using the encoded products."
A control experiment that would distinguish these two readings is straightforward to design and is not part of standard virology workflow. The fact that the distinction is not tested is part of what the audit documents.
What the critic must concede: that the experimental specificity of reverse-genetics workflows is gene-expression specificity, and that the leap from gene-expression specificity to organism replication is added by interpretation rather than established by the experimental design.
Objection Four: "Modern density gradients cleanly separate virions from exosomes."
The objection is that whatever the historical and interpretive problems with the earlier work, modern ultracentrifugation with iodixanol or sucrose gradients produces discrete bands of virion-rich and exosome-rich fractions, and the institutional separation is therefore methodologically clean.
The most informative response is to audit a recent paper that explicitly claims clean separation using modern methods. The CVB3 paper by Fu, Xiong, and colleagues (PLOS Pathogens, 2023) is well-suited because it post-dates the modern exosome literature, explicitly addresses the virion-exosome distinction, uses standardized methodology, and is written within the institutional frframework.
The paper's own data contradicts the clean-separation claim at four specific points.
First, the exosomal marker CD9 is documented in Western blot data (Figures 3G–H) as predominant in fractions F5–F7, overlapping the fractions F8–F10 where the viral capsid protein VP1 peaks. The marker is not confined to a low-density band. It co-occurs with the virion marker across the gradient. There is no clean separation visible in the marker distribution.
Second, the paper's own text states: "a portion of virions was found floating on an iodixanol cushion in the fractions containing exosomes (F6–F7), suggesting that these virions had been sedimented with exosomes." The supposedly distinct virion population is sedimenting with the exosomes, in the paper's own admission, in the paper's own prose.
Third, RT-PCR of the high-density "virion" fractions reported "no detectable complete CVB3 RNA sequences" — only "several fragments of viral RNA." The fractions claimed to contain intact replication-competent virions contain no complete genomes. By the institutional model's own definition, the fractions cannot contain intact virions, because intact virions require complete packaged genomes.
Fourth, the EM imaging in Figure 3I is captioned by the authors themselves as "exosomes-CVB3 attachment." The supposedly discrete viral particles are imaged as attached to exosomes, not as a separate free population. The follow-up immunomagnetic separation using antibodies against viral capsid proteins also failed to produce a clean virion population, because the materials are physically bound to one another.
The clean-separation claim is contradicted in the paper's own figures, prose, and captions. The critic defending modern density gradients as resolving the audit problem must defend the institutional summary of this paper against the paper's own data. The defense is unavailable because the data is in the paper, in publicly available figures, in the authors' own words.
What the critic must concede: that the institutional clean-separation claim, audited against the primary literature it relies on, is contradicted by the data in the very papers cited to support it.
Objection Five: "You are an isolated dissenter. The mainstream literature does not support these readings."
This objection is the consensus appeal. The audit findings are heterodox; the institutional position is the mainstream; therefore the audit findings are wrong.
The objection is structural rather than empirical, but it deserves a structural response. The five primary papers anchoring §5.6.S — Chahar 2018 (RSV), Wang 2018 (PRRSV), Ye 2020 (ALV-J), Xia 2023 (SARS-CoV-2), and Fu and Xiong 2023 (CVB3) — are all mainstream peer-reviewed institutional literature. They are not heterodox sources. They were written by institutional researchers, published in institutional journals, and intended to support the institutional model. The audit reads them at the physical-time layer of their own protocols and finds that the data they contain documents stressed-cell exosome biology across five virus families. The institutional framing is layered on top of the data, not derived from it.
The audit is not asking the reader to accept a heterodox source against the mainstream literature. It is asking the reader to read the mainstream literature for what its own protocols document. The pattern it produces — stressed cells release fragmented cellular cargo in exosomes; receiving cells respond with stress and inflammatory signaling; nothing replicates; no discrete organism is required to explain the observations — is consistent across five papers, five virus families, and five independent research groups. It is the institutional literature itself producing the audit finding when its data is read without the entity overlay.
The consensus appeal collapses on its own terms when the consensus literature is read directly. Either the data the consensus papers contain supports the discrete-organism model, in which case the data should be defensible against the audit point-by-point, or the data does not support the model, in which case the consensus appeal is doing the work of asserting what the data does not show.
What the critic must concede: that the audit is reading mainstream literature at the physical-time layer of its own protocols, and that the consensus appeal cannot substitute for engagement with what the data in those papers actually documents.
Objection Six: "Even if some details are contested, the overall framework of modern virology is supported by overwhelming evidence."
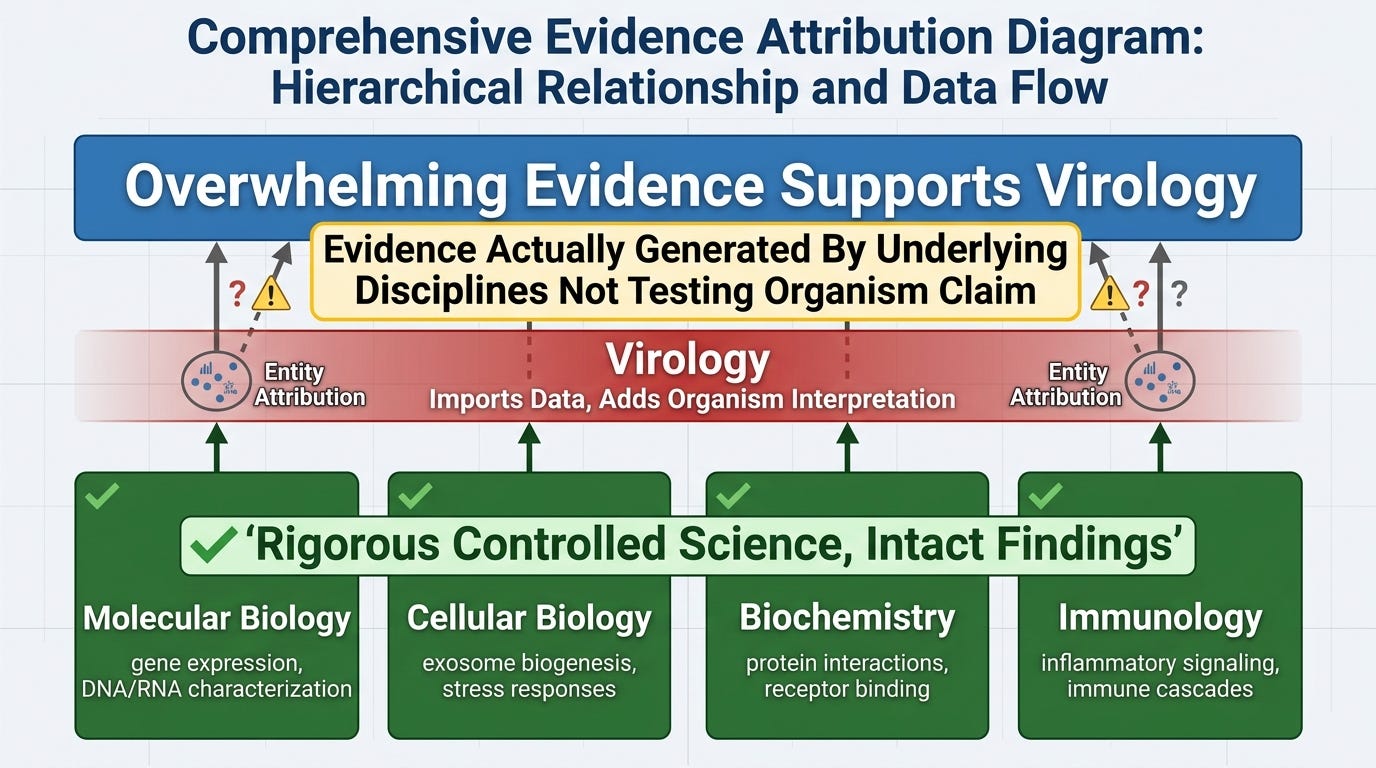
This is the totalizing defense. Individual experiments may have problems; the overall edifice is too well-supported to fall.
This objection commits the inverse of the audit's structural finding. The audit documents that virology is not an independent science generating its own empirical findings. It is a narrative layer applied to the findings of molecular biology, cellular biology, biochemistry, and immunology. The "overwhelming evidence" the critic invokes is, on inspection, the evidence those underlying disciplines have generated — gene expression studies, protein characterization, cellular response data, biochemical interaction measurements, immunological signaling cascades.
That evidence is real. It is rigorous. It is generated by researchers running controls and following scientific method to the highest standards. None of it is contested by the audit. What the audit contests is the relabeling layer that takes the molecular and cellular biology and reframes it as evidence for a discrete external organism. The molecular biologists characterizing gene expression are not virologists making organism claims. The cell biologists characterizing exosome biogenesis are not virologists making organism claims. The biochemists characterizing receptor interactions are not virologists making organism claims. Virology imports their work and adds the entity attribution.
When the relabeling is removed, what remains is the underlying disciplines doing rigorous science with intact findings. The "overwhelming evidence" supports the underlying disciplines. It does not, on inspection, support the entity claim virology adds on top, because the underlying disciplines were not designed to test that claim and the experiments they ran do not require it.
What the critic must concede: that the overwhelming evidence cited as supporting virology is the evidence generated by molecular biology, cellular biology, biochemistry, and immunology, and that the entity attribution virology adds is not what that evidence was generated to establish.
The pattern across all six objections
Each objection, examined honestly, requires the institutional defender to concede a structural element of the audit finding. The positive-control objection concedes that the institutional reference standard does not exist. The Chahar reading concedes that the institutional framing is added on top of the cellular biology rather than derived from it. The reverse-genetics objection concedes that experimental specificity is gene-expression specificity, not organism specificity. The density-gradient objection concedes that the cited primary literature contradicts the clean-separation claim. The consensus appeal concedes that the consensus literature, read directly, produces the audit finding. The totalizing defense concedes that the evidence is generated by underlying disciplines whose findings the audit does not contest.
The objections do not save the institutional model. They reveal that the model is held in place by the institutional vocabulary applied to the underlying science, not by the science itself. The audit is not asking the reader to reject science. It is asking the reader to read the science directly, without the relabeling layer, and observe what the data documents on its own terms.
The pleomorphism admission
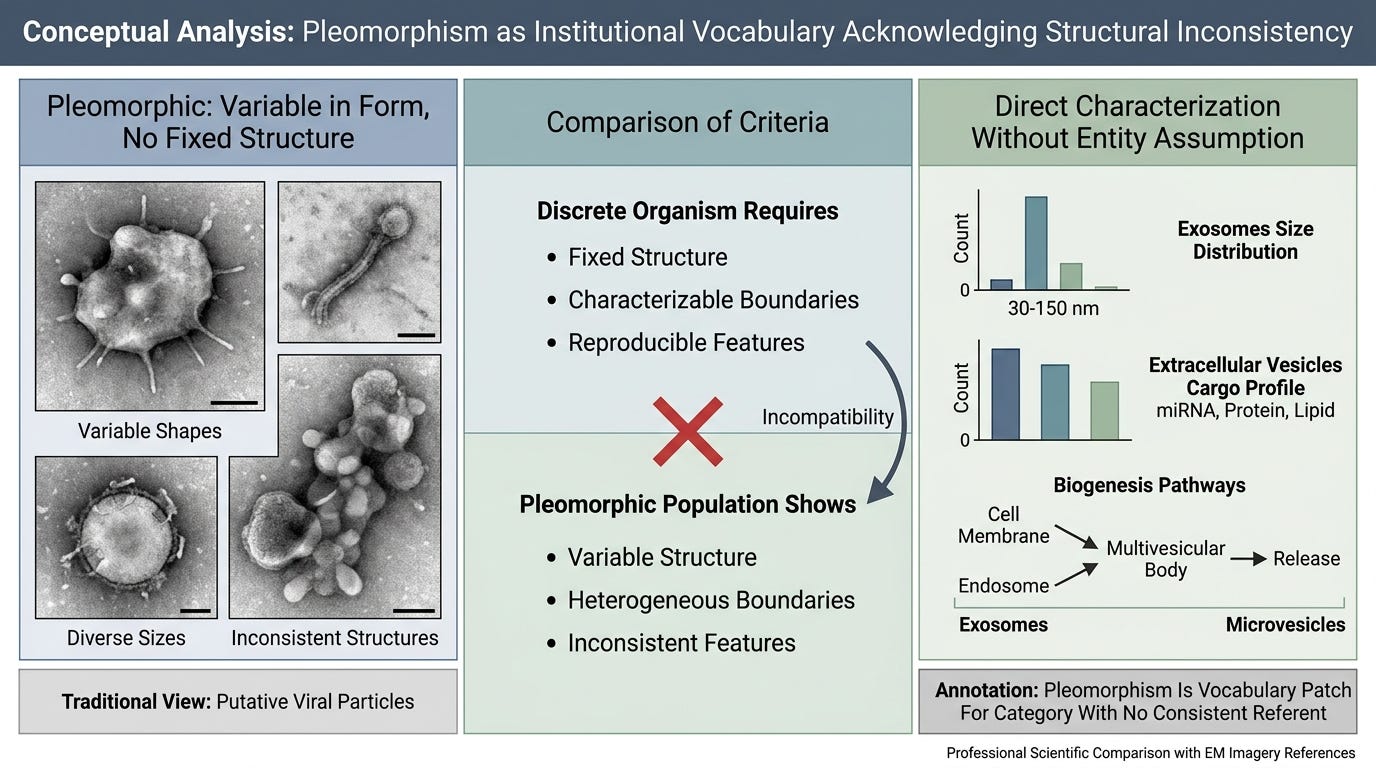
There is one final observation worth making, which the institutional literature has placed in its own vocabulary. Modern virology refers to viral particles as pleomorphic — variable in shape, size, and structural features. The term is used routinely in electron microscopy descriptions, in standard textbooks, in the characterization of major virus families.
A pleomorphic entity has no fixed structure. An entity without fixed structure does not have the properties an organism claim requires. Discrete identity, characterizable boundaries, reproducible structural features — these are the criteria that distinguish an organism from a heterogeneous population of cellular debris. Pleomorphism describes the absence of these criteria.
Cellular biology does not need the term "pleomorphic" to characterize exosomes or extracellular vesicles. It measures their size distributions, cargo profiles, biogenesis pathways, and surface markers directly. The term becomes necessary only when a population must be labeled as a single entity despite lacking the structural consistency a single entity would have. Pleomorphism is the vocabulary patch for a category that has no consistent referent.
The institutional adoption of "pleomorphic" as a routine descriptor is the field acknowledging, in its own vocabulary, that the structures it labels as viruses do not have the morphological consistency a discrete organism would have. The label is doing the work of holding together a heterogeneous population that direct characterization would resolve into the cellular biology it actually is.
Closing
The Phase 1 paper made one argument. This response document addressed the criticisms that argument predictably attracts. None of the criticisms refutes the audit. Each, examined on its own terms, requires the institutional defender to concede a structural element of the audit finding.
What is real in the evidence base of modern virology is the molecular biology, cellular biology, biochemistry, and immunology that generates the data. That work is rigorous, properly controlled, and produces hard data that can be independently verified. The audit does not contest any of it.
What the audit contests is the layer of entity attribution that virology, as a discipline, applies on top of that underlying work. The label "virus" is doing the work the physical-isolation of a discrete entity never did. Every downstream technique inherits the label and operates within it. The institutional model is held in place not by the science but by the vocabulary applied to the science.
When the vocabulary is removed and the data is read directly, what remains is the cellular and molecular biology that the underlying disciplines characterize accurately, doing the work they have always done. The invisible organism is the label. The cellular biology is the reality.
The audit is complete to the extent that the institutional model has been falsified at the physical-evidence layer. The terrain reading — the older observational account of illness as the body's response to physical and environmental conditions — is what is left when the nineteenth-century overlay is examined for whether it ever cleared its own evidentiary standard. It did not.
That is the finding. The objections do not change it. They confirm it in different vocabulary.