
Reverse Genetics Decoded: How Cellular Detoxification Got Mislabeled as Viral Replication
The Identification That Was Never Made

Under the Terrain Framework
Reverse Genetics is not evidence of an autonomous virus—it is the ultimate demonstration of how a living cell reacts to an artificial, laboratory-engineered genetic toxin in an environment deliberately designed to kill it.
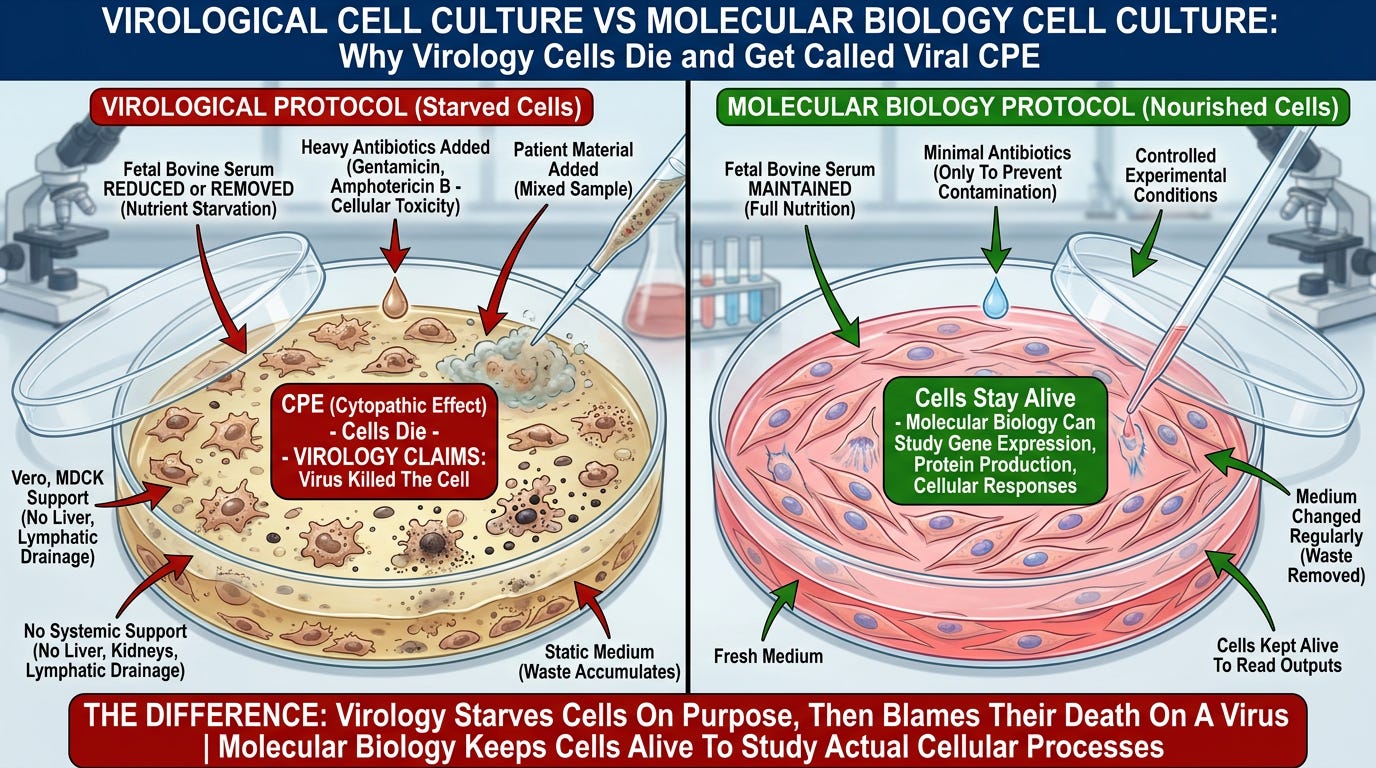
In this model, the synthesized genetic sequence is not a blueprint for an independent organism. Instead, it is an artificial chemical irritant introduced into starved, antibiotic-poisoned, isolated cell cultures that are already dying from the laboratory protocol itself.
The critical observation: Virology conducts reverse genetics in cell cultures that have been:
- Starved of nutrients (fetal bovine serum reduced or removed)
- Poisoned with heavy antibiotics (gentamicin, amphotericin B)
- Severed from systemic support (no liver, kidneys, lymphatic drainage to clear waste)
- Forced to accumulate their own metabolic waste (static medium, no waste removal)
In contrast, molecular biology keeps cells alive and healthy by maintaining full nutrition, minimal antibiotics, and regularly refreshed medium—because living cells are required to study actual cellular processes.
Virology’s methodology ensures cells will die, then attributes that death to the synthetic sequence it introduced, calling it “viral replication.”
The sequencing fraud: When virologists sequence the particles released from these dying cells, they claim to have found “viral RNA.” But what they actually detected are fragments released during the cellular breakdown cascade—the cell’s apoptotic death and waste-elimination process. The synthetic sequence was broken into fragments by the dying cell for exosomal packaging and elimination. At no point during the cell-culture experiment did a single full viral genome get copied, reproduced intact, and released as a complete unit. And crucially, virologists have never extracted a whole genome from inside a single particle—they always sequence millions of short fragments and computationally assemble them, even after forcing the synthetic construct into cells.
This article presents the complete physical-evidence audit of what virology calls “viral replication” and “viral genomes,” demonstrating that:
1. The sequences detected by PCR are real but are cellular stress-response transcripts (in natural exposure) or the fragmented synthetic construct released during cellular breakdown (in lab reverse genetics)—neither proves organism existence
2. The “viral genome” is a computational construct assembled from millions of short fragments released by dying cells, never isolated as a complete molecule from a single particle—even after reverse genetics
3. Reverse genetics proves cells fragment and eliminate synthetic inputs during apoptotic death, not organism replication
4. The RNA sequences fall into two categories:
- Natural exposure: Toxin-response hybrids generated through proteasome breakdown, MHC processing, and reverse transcription of environmental insults
- Lab reverse genetics: Fragments of the synthetic construct you forcibly introduced, broken up by dying cells during their elimination cascade and apoptotic collapse
- Neither proves organism existence
5. Sequence convergence across patients (in natural outbreaks) reflects shared environmental exposure, not organism transmission
6. The laboratory protocol itself kills cells—a fact demonstrated by Dr. Stefan Lanka’s control experiments, where cells die identically without any patient material added, releasing identical particles
What virology attributes to an invisible pathogen is entirely explicable through:
- In nature: Documented cellular stress-response biology (transposable-element activation, toxin breakdown, environmental-insult signatures)
- In the lab: Cell death from starvation + toxicity + forced processing of synthetic inputs → cellular breakdown cascade → fragmentation and elimination of the introduced sequence via apoptosis and exosomal packaging
The particles released are cellular waste, not organisms. The RNA detected is breakdown debris, not a genome. No discrete organism required in either scenario.
This captures the complete fraud of viral cell culture: “They force cells to die, collect the breakdown debris, sequence the fragments, and call it “viral RNA” - when it’s actually just the cell’s elimination of the toxic input they introduced!”
SECTION ONE: The Reverse Genetics Detoxification Cascade
Reverse Genetics Detoxification Cascade
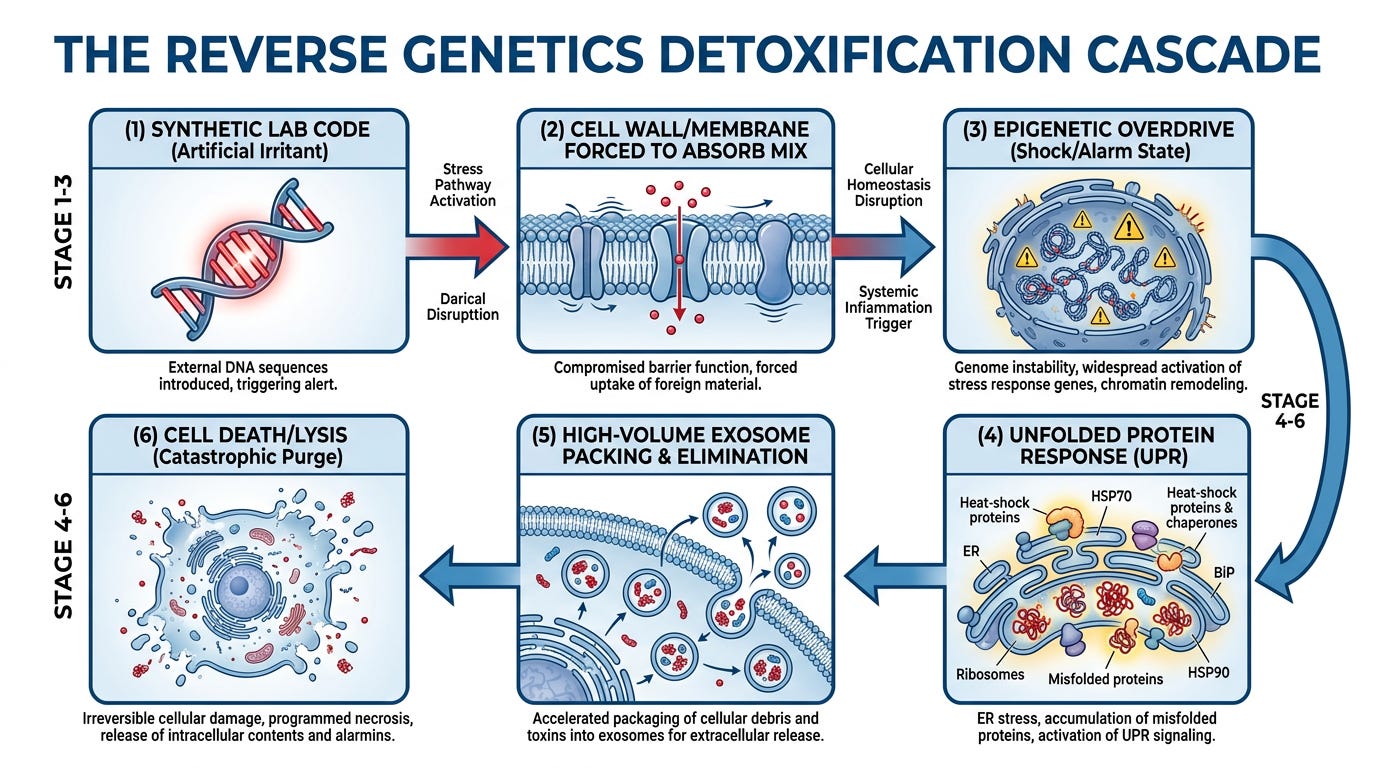
The Complete Pathway Unfolds in Six Stages:
Stage 1: Introduction of Synthetic Material
In a laboratory, scientists chemically manufacture a specific genetic sequence (DNA or RNA) from raw chemicals or a computer text file. Because this sequence has been engineered to match specific cellular receptors, it acts as a hyper-targeted chemical key.
When dropped onto cells in a dish, the cells are forced to absorb this foreign, non-native material through their cell membranes—not because a predator invaded, but because the synthetic code was chemically designed to exploit the cell's natural uptake pathways.
Stage 2: The Epigenetic Shock State
Epigenetic Shock
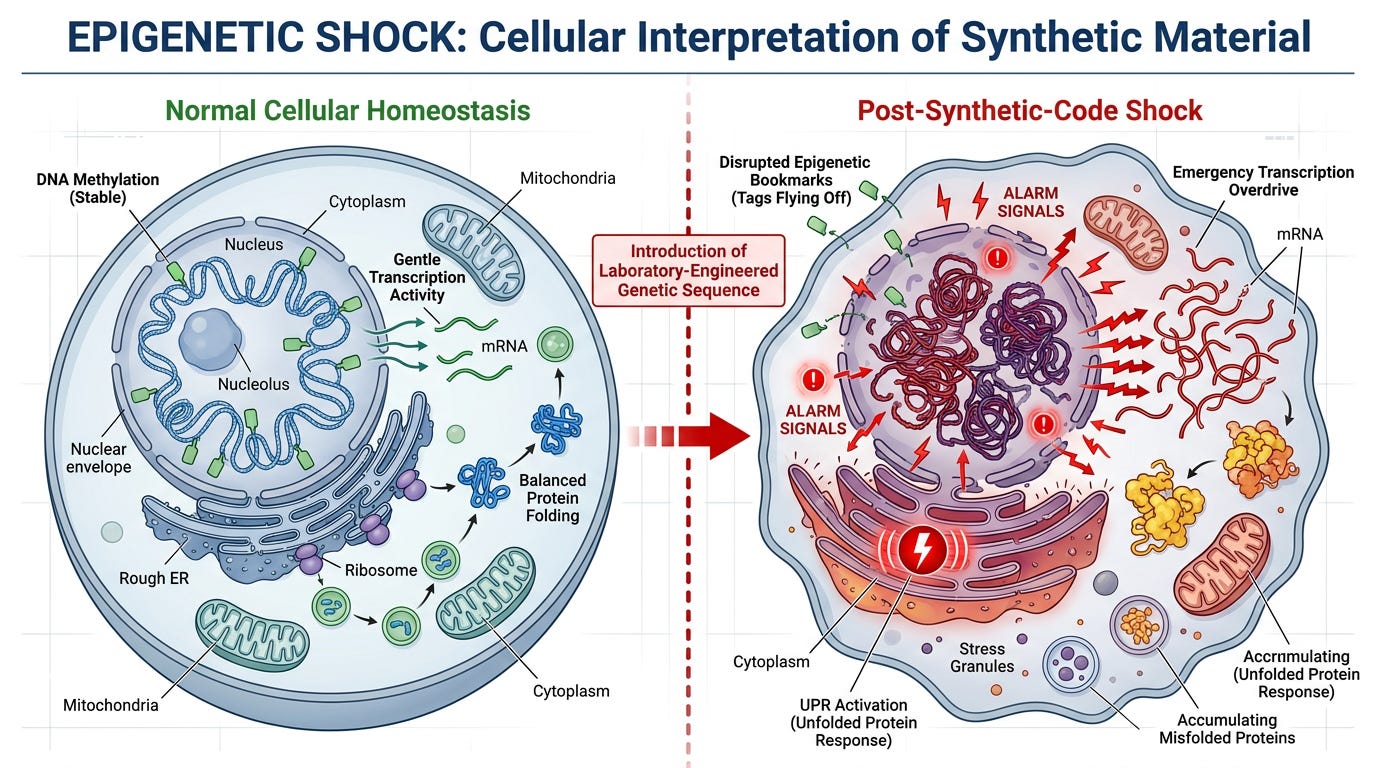
The moment this synthetic, foreign genetic code enters the cytoplasm, it acts as a severe internal stressor. It drastically disrupts the cell's homeostatic balance and throws its epigenetic bookmarks into chaos.
The cell's nucleus reads this jarring influx of foreign code and interprets it as a state of extreme cellular trauma. Chemical methyl tags that normally regulate gene expression begin destabilizing. Emergency transcription pathways activate. The cell enters full alarm mode.
Key Point: This is not infection—this is chemical shock. The cell is responding to the synthetic sequence exactly as it would respond to any severe environmental toxin: heavy metals, chemical burns, or radiation damage.
Stage 3: The Unfolded Protein Response (UPR)
To defend itself against this synthetic genetic overload, the host cell activates an emergency survival mechanism known as the Unfolded Protein Response (UPR).
- The cell stops its normal, everyday biological functions
- It marshals all of its internal energy to deal with the foreign material
- Because the synthetic code forces the cell's ribosomes to manufacture unfamiliar, highly irritating protein structures, the cell immediately treats these proteins as hazardous metabolic waste
The endoplasmic reticulum (ER) becomes overwhelmed with misfolded proteins. Heat-shock proteins are recruited en masse. The cell is now in a state of metabolic emergency.
Stage 4: High-Volume Exosomal Elimination
Exosomal Elimination
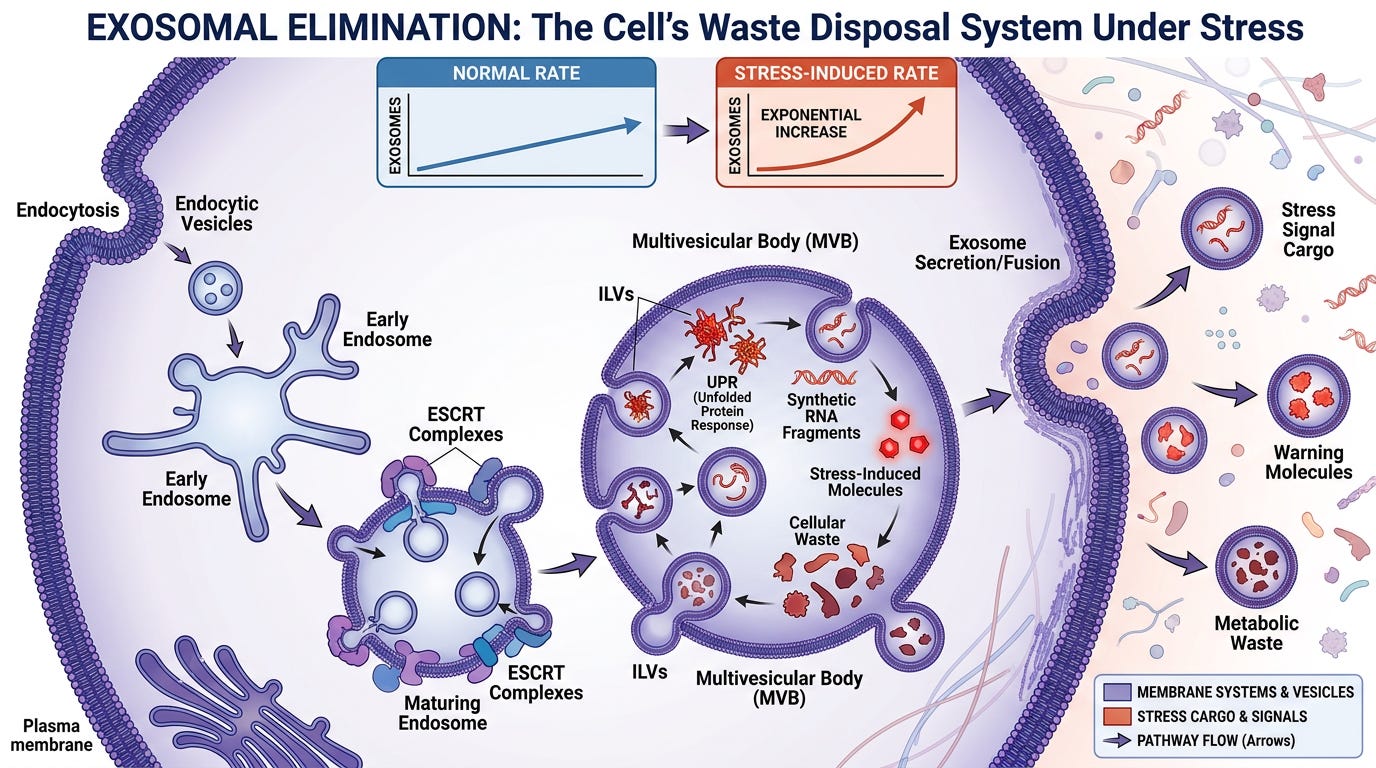
The cell must eliminate this artificial waste to survive. It activates its internal recycling and disposal networks, utilizing its native MHC domains and cellular membranes to package the toxic laboratory code and its matching proteins into microscopic waste disposal sacs.
Virology labels these as "viruses."
The Terrain Framework recognizes them as stress-induced exosomes.
The cell begins pumping these vesicles out at an exponential rate to purge the synthetic irritant from its system and warn neighboring cells of the localized toxicity. This is not replication—this is high-volume waste elimination.
Stage 5: Catastrophic Cellular Collapse (Lysis)
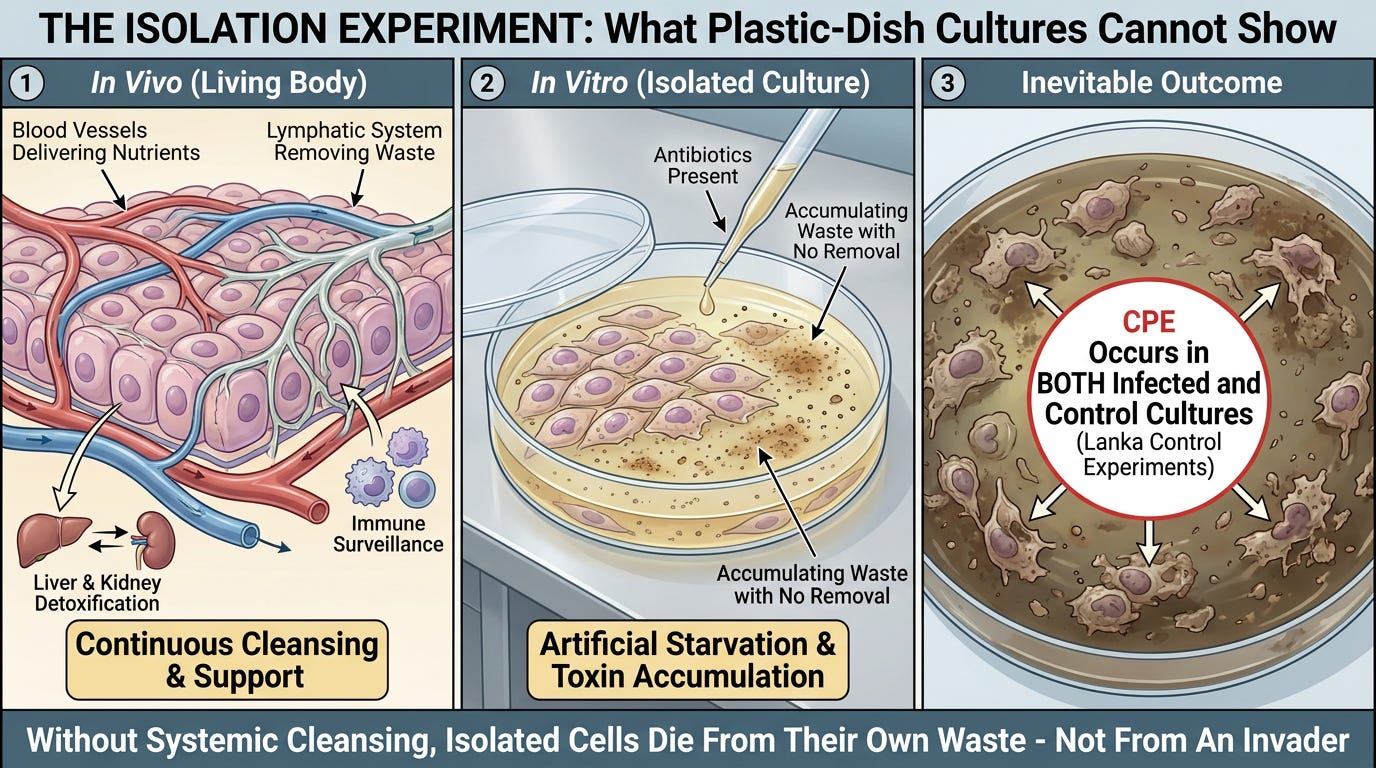
Isolation Experiment Problem
Because cell cultures in a laboratory plastic dish are completely isolated—lacking a lymphatic system, a liver, or kidneys to flush the waste away—the cells quickly drown in their own manufactured elimination debris.
When the cells reach their absolute physical limit of stress and toxin accumulation, they undergo apoptosis or lysis (they burst and die).
Virology points to this cell death and claims: "The virus killed the cell."
The Terrain Framework proves the exact opposite: The cell worked itself to death trying to process, package, and eliminate a massive, unnatural influx of synthetic laboratory material.
SECTION TWO: Who Does The Work? The Cell As Active Factory
The Cell As Active Factory
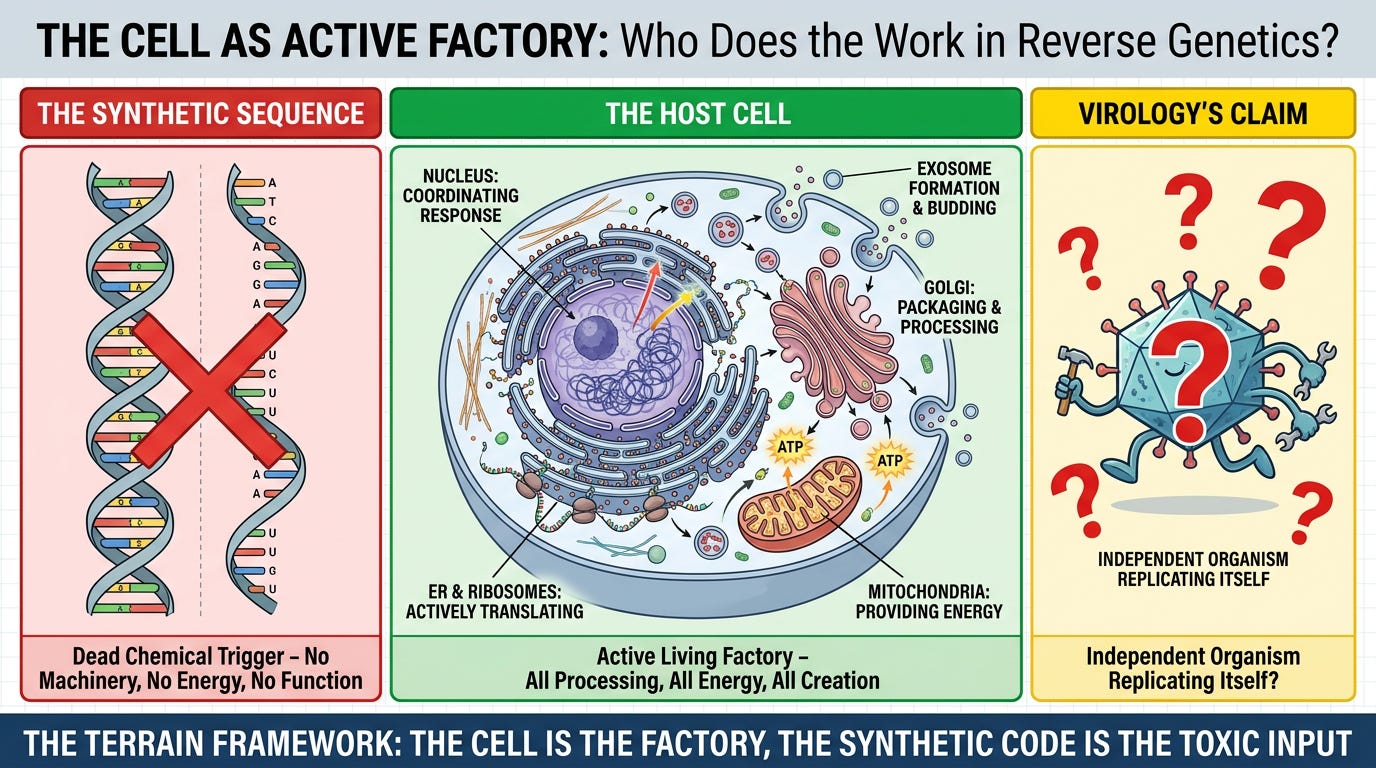
A critical question exposes the entire framework:
In a Reverse Genetics experiment, which entity possesses the biological machinery to transcribe, translate, process, package, and eliminate?
The Synthetic Sequence:
- No ribosomes
- No energy source (no ATP)
- No protein-folding machinery
- No membrane systems
- No organelles
- Cannot replicate itself outside a living cell
Status: Dead chemical trigger. Passive information. Requires a host factory to do anything.
The Host Cell:
- Active ribosomes translating RNA → protein
- Endoplasmic reticulum folding proteins
- Golgi apparatus packaging cargo
- Mitochondria supplying energy (ATP)
- ESCRT complexes forming exosomes
- Plasma membrane budding vesicles
- All processing, all energy, all creation
Status: The entire factory. The living terrain performing every observable function.
Virology's Claim:
- The synthetic sequence is an "independent organism"
- It "hijacks" the cell
- It "replicates itself"
- It "produces progeny virions"
Terrain Response: The cell IS the factory. The synthetic sequence is merely the toxic input that forces the factory into emergency overdrive. Calling this "viral replication" is like calling a car factory's recall of defective parts "the defect replicating itself."
SECTION THREE: Cytopathic Effect (CPE) — Two Interpretations
Cytopathic Effect
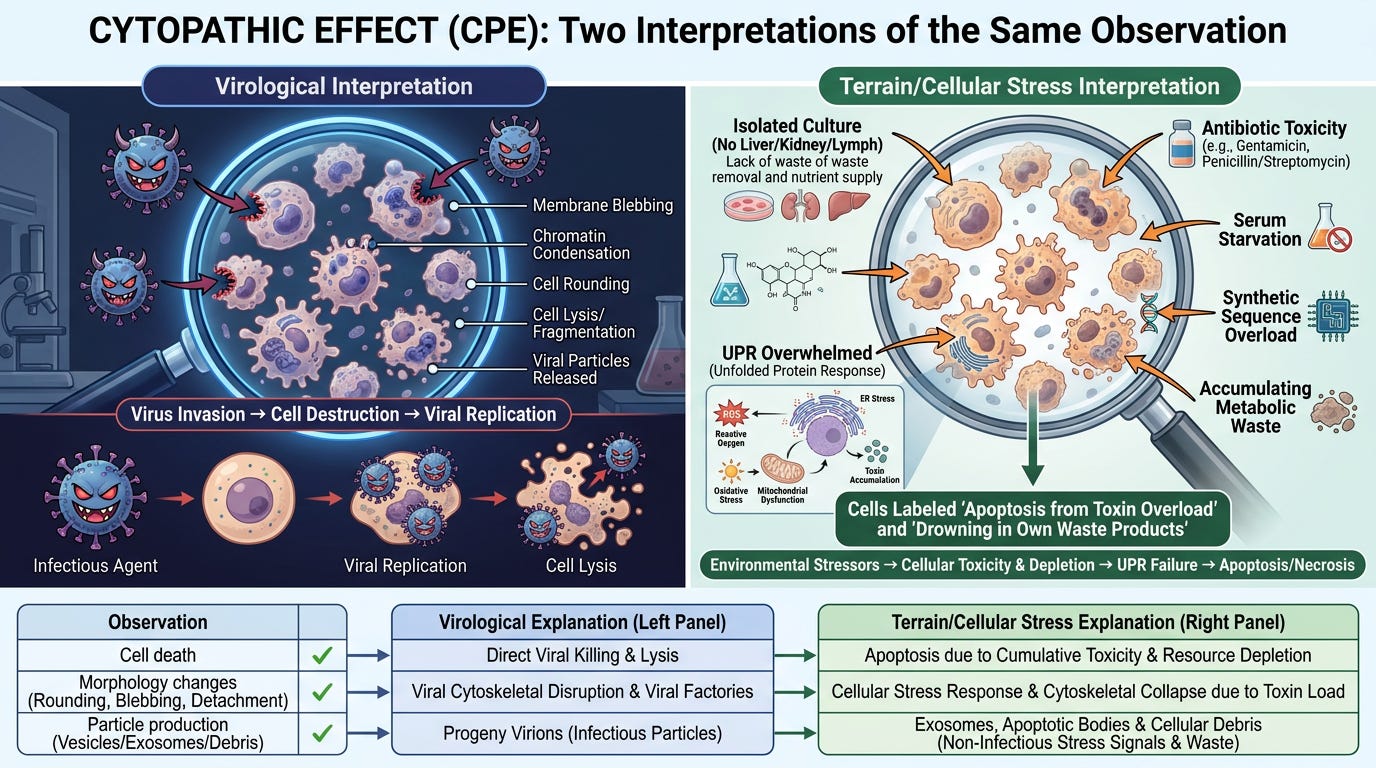
When cells die in culture after exposure to a "virus," mainstream virology interprets this as proof of viral destruction. The Terrain Framework offers a radically different—and biochemically precise—explanation.
Virological Interpretation:
1. Virus invades cell
2. Virus replicates inside cell
3. Virus destroys cell from within
4. Cell death = proof of viral pathogenicity
Terrain/Cellular Stress Interpretation:
1. Isolated culture environment: Cells severed from liver, kidneys, lymphatic drainage
2. Chemical toxicity: Antibiotics (gentamicin, amphotericin B) added to prevent bacterial "contamination"
3. Serum starvation: Fetal bovine serum reduced or eliminated
4. Synthetic sequence overload: Foreign genetic material forces UPR activation
5. Accumulating metabolic waste: No systemic clearance; exosomes and debris accumulate in static medium
6. Cellular apoptosis: Cells drown in their own waste products
Both observations produce identical microscopic images of dying cells. The difference is the root cause assigned.
The Lanka Control Experiments
Dr. Stefan Lanka demonstrated that CPE occurs in control cultures that receive no patient material whatsoever—only the starvation protocol and chemical additives. This proves CPE is an artifact of laboratory conditions, not evidence of an invading organism.
SECTION FOUR: The "Viral Genome" Objection — Two Completely Different Scenarios
The Most Common Objection:
"But they've sequenced the full viral genome — 30,000 base pairs for SARS-CoV-2, 7,400 base pairs for CVB3. If the virus doesn't exist, where did that sequence come from? Reverse genetics uses that exact sequence and produces the exact same effects. Doesn't that prove the genome is real and the virus exists?"
This objection conflates two completely different scenarios:
SCENARIO A: Natural environmental exposure (what happens in real-world outbreaks)
SCENARIO B: Forced synthetic input in a petri dish (what happens in reverse genetics labs)
Neither scenario demonstrates organism existence. Here's why:
SCENARIO A: Natural Environmental Exposure (Real-World Outbreaks)
Computational Assembly vs Physical Isolation
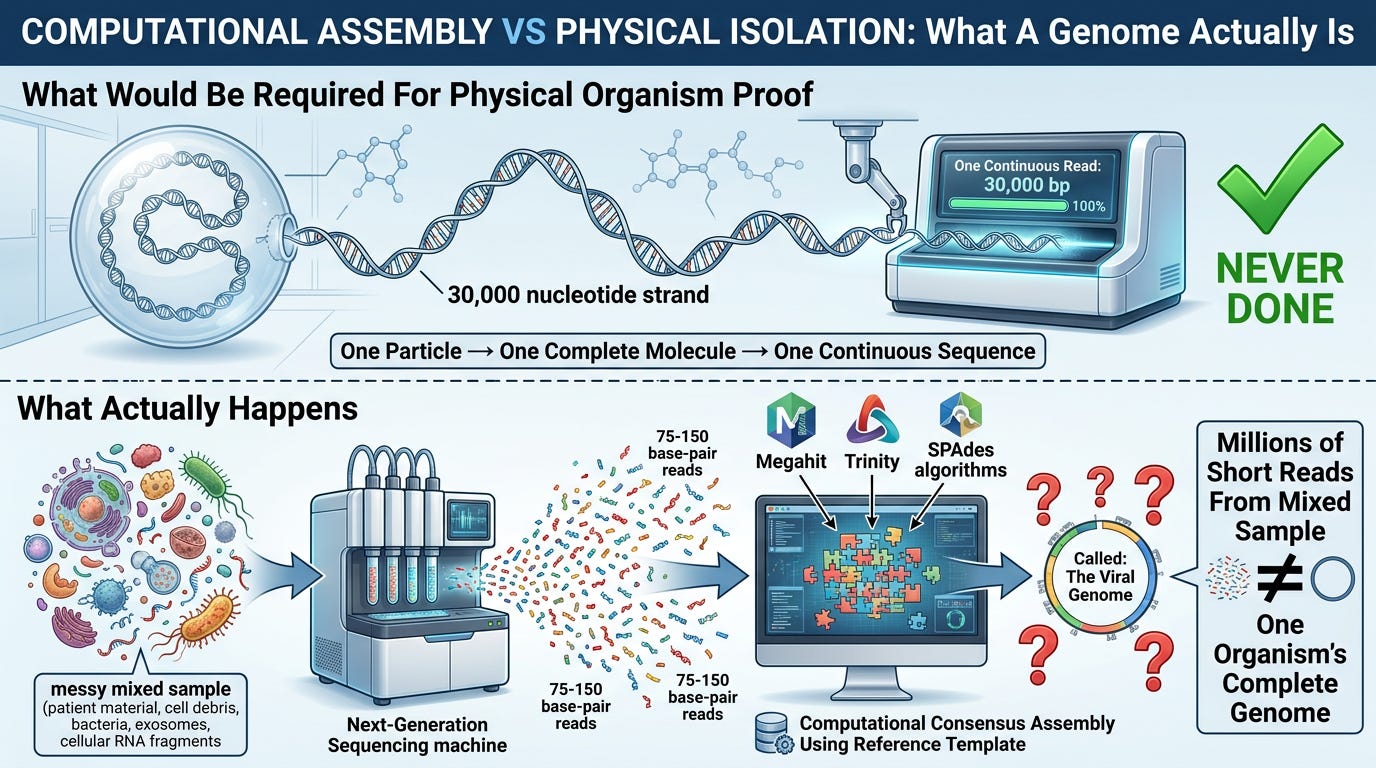
What Actually Happens:
1. Environmental Insult Occurs:
- Population exposed to pollution spike, EMF rollout, chemical contamination, mass vaccination campaign, nutritional deficiency, or toxic environmental event
2. Cellular Stress-Response Activates:
- Cells transcribe stress-response genes (interferon pathways, heat-shock proteins, inflammatory markers)
- Transposable elements activate (LINE-1, Alu, HERV sequences from "junk DNA")
- Toxin-breakdown signatures generated through MHC processing and reverse transcription
3. RNA Fragments Produced:
- These are short fragments (hundreds to thousands of bases) from cellular stress-response pathways
- They are endogenous human sequences—from the patient's own genome responding to environmental stress
- They tell you what poisoned the patient (environmental-insult signatures)
4. Sample Collection and Processing:
- Collect mixed sample from sick patients (contains human cells, bacteria, exosomes, cellular debris, mitochondrial RNA)
- Filter through 0.22 μm filter (removes bacteria but leaves mixed nanoparticle soup—exosomes, cell fragments, protein aggregates)
- Extract total RNA (millions of short fragments from all sources mixed together)
5. Computational Assembly:
- Next-Generation Sequencing produces millions of short reads (75-150 base pairs each)
- Bioinformatics software (Megahit, Trinity, SPAdes) assembles fragments using:
- Overlap algorithms
- Reference templates from databases (existing "viral genomes")
- Consensus building (majority rule for ambiguous positions)
- Gap-filling (inferring missing sections)
6. Output: The "Viral Genome"
- A 30,000 bp consensus sequence is generated
- This sequence never existed as a single intact molecule in any particle
- It is a computational construct—a software-generated assembly from mixed-sample fragments
- It is labeled "viral genome" and uploaded to databases
What This Actually Proves:
✓ Patients were exposed to environmental stress
✓ Cells produced stress-response RNA fragments
✓ Those fragments are real and detectable
✓ Computational assembly can stitch them into a consensus sequence
✗ Does NOT prove a discrete organism exists
✗ Does NOT prove the sequence came from a single particle
✗ Does NOT prove organism replication
The "genome" is a labeled assembly of cellular stress-response transcripts—environmental-insult signatures, NOT an organism's genetic code.
SCENARIO B: Reverse Genetics In A Petri Dish (Laboratory Validation Attempt)
Genome Circularity Problem
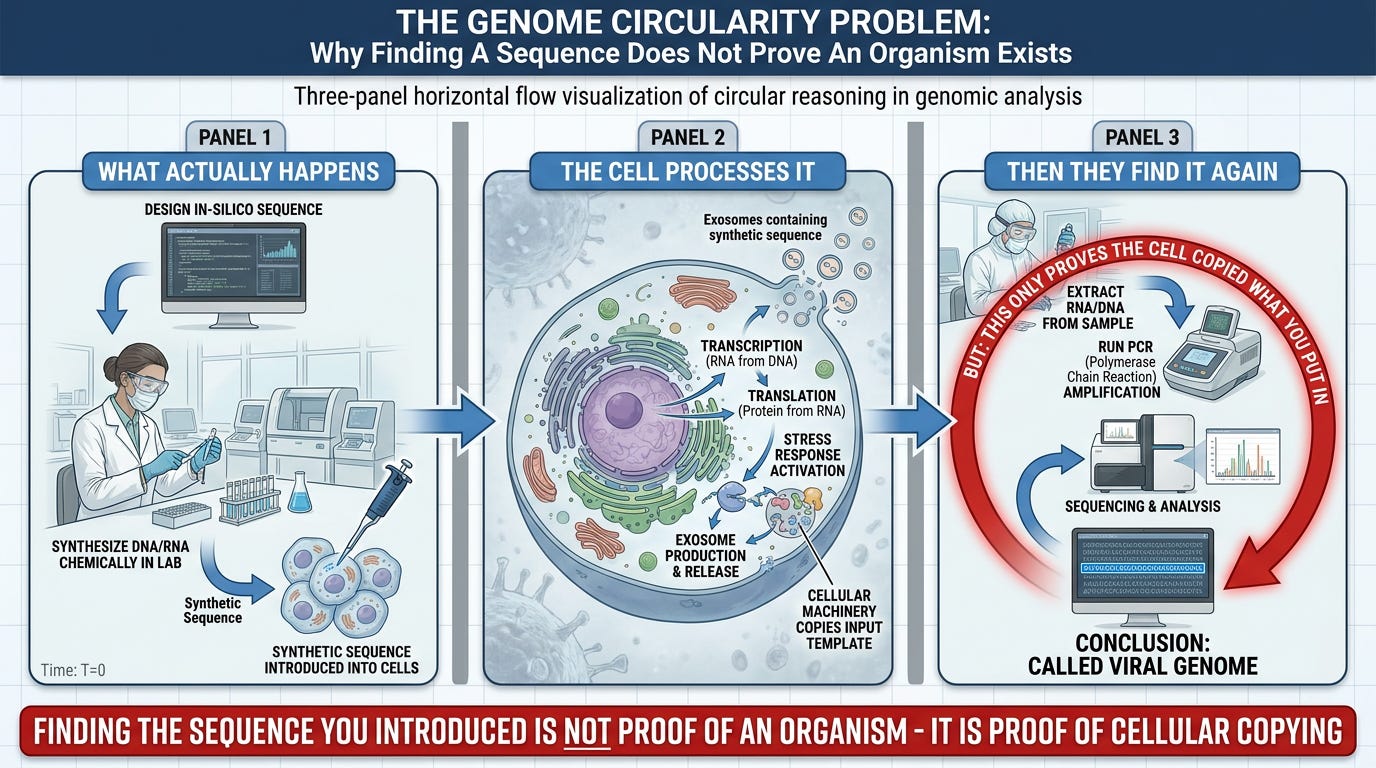
What Actually Happens:
STAGE 1: Create The Fiction
1. Take the computationally-assembled "genome" from Scenario A
2. This consensus sequence (that never existed as an intact molecule in nature) becomes the "reference"
STAGE 2: Synthesize The Fiction
3. Chemically manufacture the entire 30,000 bp sequence base-by-base in a laboratory
4. This is a wholly artificial construct—it was never extracted from a particle, never proven to belong to an organism
5. Introduce this synthetic sequence into cell cultures that have been:
- Starved of nutrients (serum reduction)
- Poisoned with antibiotics (gentamicin, amphotericin B)
- Isolated from systemic support (no liver, kidneys, lymphatic drainage)
STAGE 3: Cellular Processing
6. Cells absorb the synthetic sequence (via lipid nanoparticles, transfection, electroporation)
7. Cells transcribe and translate it (forced processing using their own machinery)
8. Cells undergo stress-response cascade (UPR, exosome production, inflammation)
9. Cells copy the synthetic sequence during division (DNA/RNA polymerase faithfully replicates the input)
10. Cells break the sequence into fragments for exosomal packaging and waste elimination
STAGE 4: "Validation" Sequencing
11. Extract RNA from cell culture
12. Even after introducing a whole synthetic sequence, extraction yields millions of short fragments (cells have broken it up for processing and elimination)
13. Sequence using NGS (produces 75-150 bp reads)
14. Computationally assemble fragments (matching the synthetic input sequence)
15. PCR with primers designed to match the synthetic input detects it
STAGE 5: Circular Conclusion
16. "The viral genome is validated!"
What This Actually Proves:
✓ Cells can transcribe synthetic inputs
✓ Cells can copy sequences during division
✓ Cells fragment sequences for exosomal packaging
✓ PCR can detect sequences you designed primers for
✗ Does NOT prove a discrete organism exists
✗ Does NOT prove organism replication
✗ Even in the lab, no single particle is shown to contain the full 30,000 bp sequence
All you've proven is:
- You can manufacture a synthetic construct
- You can force it into starved cells
- You can detect what you forcibly introduced
- The organism assumption remains unvalidated
THE CRITICAL DISTINCTION:
| Aspect | Natural Exposure (Real World) | Reverse Genetics (Lab) |
|---|---|---|
| What enters cells | Environmental toxin (pollution, EMF, chemical, damaged proteins) | Synthetic whole-genome construct (lab-manufactured from the Scenario A fiction) |
| RNA produced | Endogenous stress-response transcripts (interferon, heat-shock, transposable elements) | Transcripts of the synthetic input + cellular stress-response to forced processing |
| What PCR detects | Toxin-breakdown signatures + human stress-response genes | The exact sequence you put in (because primers were designed to match it) |
| Organism evidence | NONE (cellular response to environmental insult) | NONE (cellular copying of synthetic input) |
| Full genome in single particle | NO (never demonstrated) | NO (still not demonstrated—still assembled from fragments) |
| What it proves | Cells respond to environmental stress with reproducible molecular signatures | Cells can copy synthetic inputs—nothing more |
The devastating observation:
In BOTH scenarios:
- No single particle is ever shown to contain the full genome
- Sequencing always produces millions of short reads
- Software always assembles the "genome" from fragments
- The organism assumption is never independently validated
The ONLY difference:
- In natural exposure, PCR detects endogenous human stress-response sequences (tells you what poisoned the patient)
- In reverse genetics, PCR detects the synthetic sequence you forcibly introduced (tells you the cell copied what you put in)
Neither proves an organism exists.
The Perfect Circular Reference:
Step 1: Outbreak occurs → cells produce stress-response RNA → computationally assemble fragments → label it "viral genome" → upload to database
Step 2: Download "genome" from database → chemically synthesize it → force into starved cells → cells copy it → PCR detects it → "Genome validated!"
This is like:
1. Finding smoke residue in burned houses
2. Chemically analyzing the residue to create a "smoke profile"
3. Synthesizing those chemicals in a lab
4. Pumping them into a new house
5. Detecting those chemicals with a tuned detector
6. Announcing: "We've proven the Smoke Organism is real!"
You haven't proven an organism. You've proven you can manufacture chemicals, introduce them, and detect them again. The circularity is perfect.
SECTION FIVE: PCR Detects Real Sequences—But They're Environmental-Insult Signatures
PCR Detects Environmental Stress Signatures
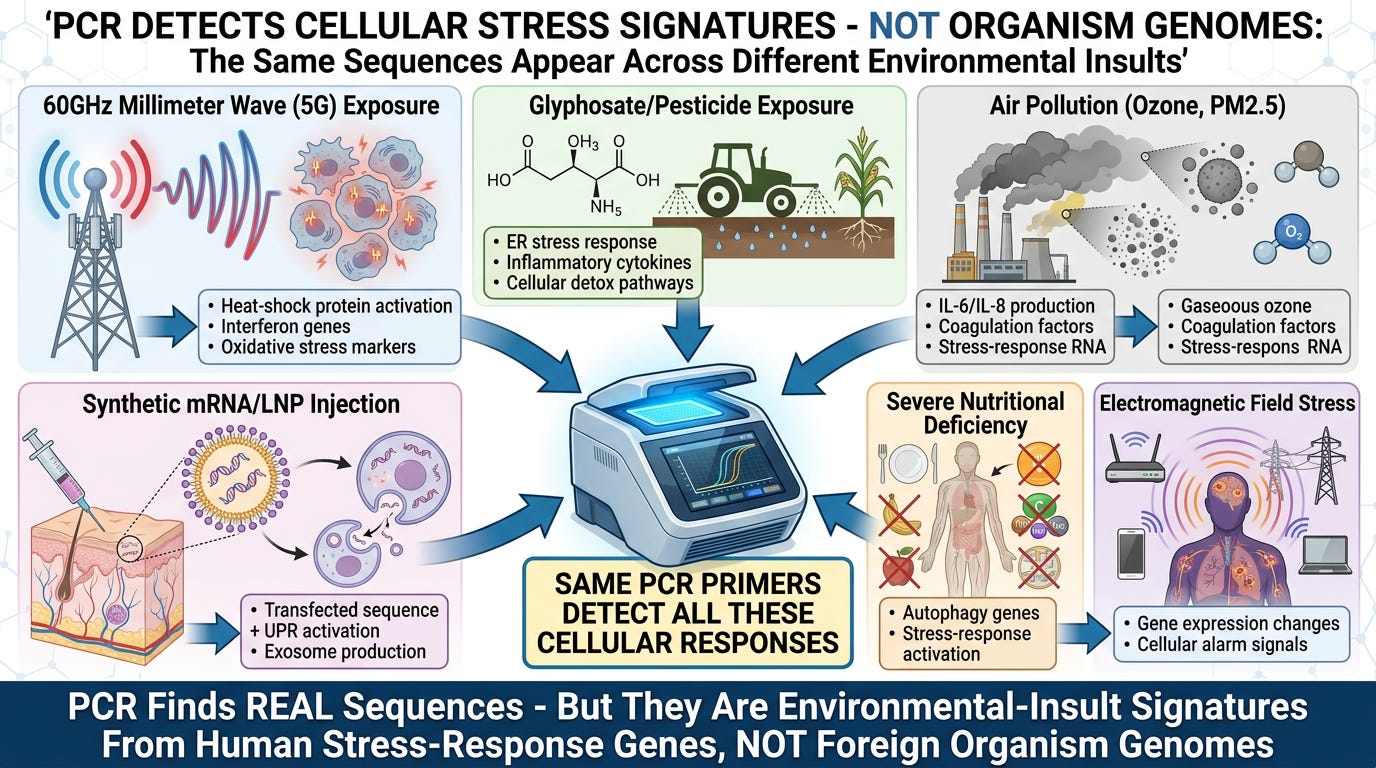
The Critical Distinction:
The audit does NOT claim PCR finds nothing at Ct 35-40.
PCR correctly detects real RNA sequences. The fraud is in how those sequences are interpreted.
Two competing interpretations:
| What PCR Finds | Virology's Interpretation | Terrain Interpretation |
|---|---|---|
| Specific RNA sequences at Ct 35-40 | "Rare viral genome from organism" | "Cellular stress-response transcript from environmental insult" |
| Interferon-stimulated genes | "Proof of viral infection" | "Endogenous stress-response to toxin, EMF, pollution" |
| Heat-shock protein mRNA | "Cell responding to virus" | "Cell responding to environmental stress" |
| Exosome-associated RNA | "Viral RNA being shed" | "Cellular waste-elimination" |
Both agree PCR found sequences. The dispute is: Are these from an organism, or from cells responding to environmental stress?
Evidence: Same Sequences Appear During Non-Infectious Environmental Exposures
Peer-reviewed research documents that specific RNA sequences appear during proven non-infectious exposures:
1. Air Pollution (Ozone, PM2.5):
- IL-6 and IL-8 elevation (inflammatory cytokines)
- Interferon-pathway activation
- Coagulation-factor changes
- Oxidative-stress markers
- Respiratory epithelial cell damage
These are the exact markers virology calls "viral infection symptoms."
2. Electromagnetic Field (EMF) Exposure (60GHz, RF-EMF):
- Heat-shock protein (HSP70, HSP90) upregulation
- Oxidative-stress gene activation
- Cellular stress-response pathway engagement
- Altered gene expression in stress-response genes
PCR primers designed to detect "viral sequences" would detect these stress-response transcripts.
3. Chemical Toxins (Glyphosate, Pesticides):
- ER-stress response activation
- Inflammatory cytokine production
- Mitochondrial dysfunction markers
- Cellular detoxification pathway upregulation
Again, these match the "viral infection" signature.
4. Synthetic mRNA/LNP Injection:
- The transfected sequence itself (by design)
- UPR activation (Unfolded Protein Response)
- Exosome production carrying sequence fragments
- Inflammatory cascade activation
- Stress-response gene transcription
PCR will detect both the transfected sequence AND the cellular stress-response to it.
Why The Same "Viral Genome" Appears Across Multiple Patients:
Virology's explanation:
"The virus spread from person to person. That's why everyone has the same genome sequence."
Terrain explanation:
"Everyone in the same geographic region was exposed to the same environmental insult (pollution event, EMF infrastructure, contaminated water, mass vaccination campaign). Their cells responded with the same stress-response gene activation. PCR detected those shared cellular signatures."
The sequences converge because:
1. Human cells have a limited repertoire of stress-response pathways (interferon system, heat-shock response, ER stress, inflammatory cascades)
2. Specific insults trigger specific pathways (EMF → heat-shock; pollution → oxidative stress + inflammation; synthetic mRNA → transfection + UPR)
3. PCR primers target these stress-response genes
4. Everyone exposed to the same insult produces the same transcripts
This explains:
- Geographic clustering of "cases" (localized pollution, regional EMF deployment, targeted vaccination campaigns)
- Temporal clustering (outbreak timing matches environmental event timing)
- Sequence similarity across patients (same insult → same cellular response → same transcripts detected)
No organism required. Just shared environmental exposure producing shared cellular stress signatures.
SECTION SIX: The "Not In Human Genome" Objection—The Toxin-Response Hybrid Mechanism
Complete Mechanism
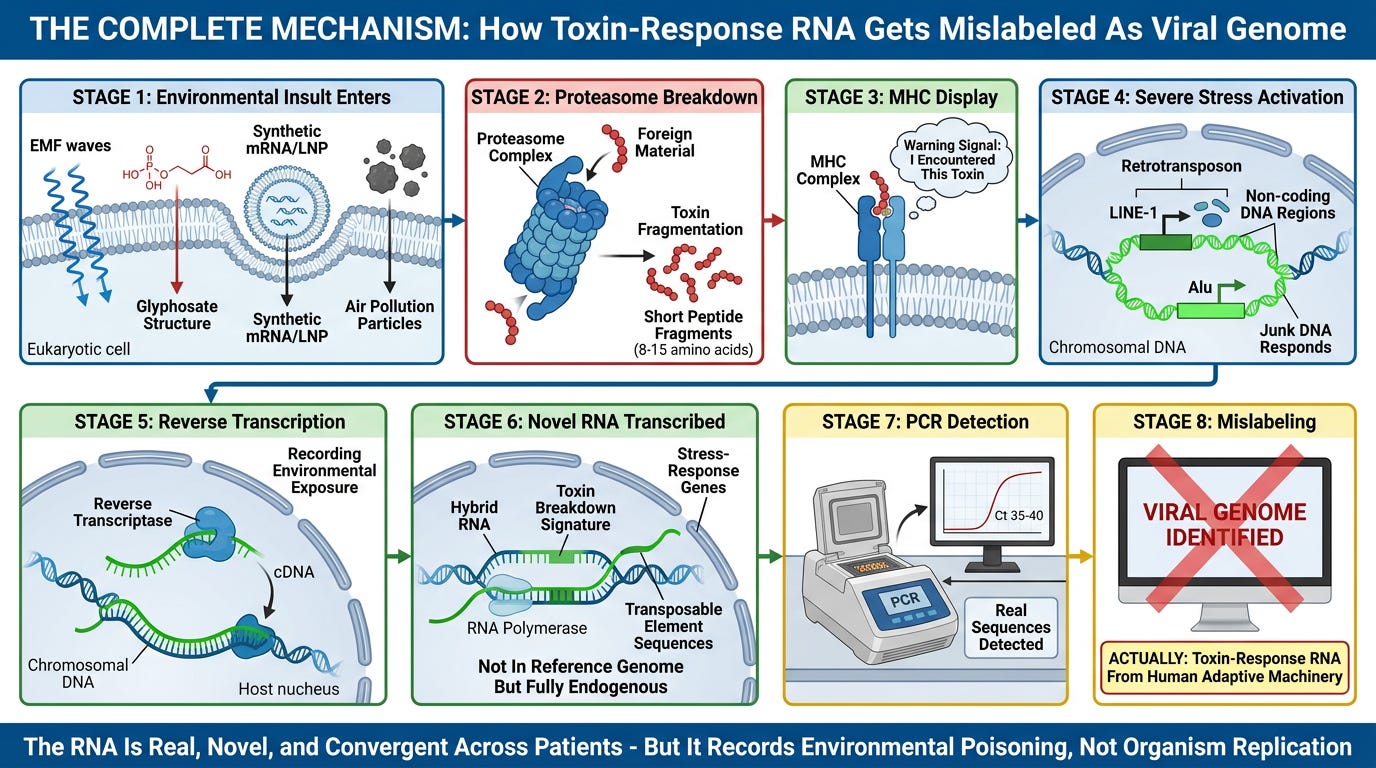
The Final Objection:
"The viral RNA sequences are NOT found in the human genome. They are novel, foreign sequences. Therefore, they must come from an external organism."
Why This Is A Category Error:
It's true you won't find the exact "viral genome" as a continuous 30,000 bp stretch in the human reference genome. But this misunderstands how cellular stress-response RNA is generated in natural environmental exposure (Scenario A).
In natural exposure, the RNA is NOT "human" in the sense of coming from protein-coding genes—it's a toxin-response hybrid sequence generated through documented cellular mechanisms.
Note: This mechanism applies to Scenario A (natural environmental exposure), NOT Scenario B (reverse genetics lab work, where you're detecting the synthetic construct you forcibly introduced).
The Complete Process (Natural Environmental Exposure):
Step 1: Foreign Toxin/Material Enters Cell
- Could be: damaged proteins from pollution, EMF-induced cellular damage, chemical toxin (glyphosate), heavy metals, pesticide residue
- The cell recognizes this as foreign, non-self material
Step 2: Proteasome Shreds The Toxin Into Peptide Fragments
- The proteasome (cellular waste-processing machinery) breaks down the foreign protein/toxin into short amino-acid chains (peptides)
- These fragments are 8-15 amino acids long
Step 3: MHC Complexes Display The Fragments
- Major Histocompatibility Complex (MHC) molecules physically bind these peptide fragments
- The MHC-peptide complex is displayed on the cell surface as a warning flag: "I encountered this toxin"
- Critical point: The peptide fragment is physically incorporated into the MHC structure—it becomes part of the antenna
Step 4: The Cell Records The Exposure (Reverse Transcription)
- Under severe stress, the cell activates LINE-1 retrotransposons (mobile genetic elements in "junk DNA")
- LINE-1 elements encode reverse transcriptase (RT)—the enzyme that converts RNA back into DNA
- The cell transcribes RNA reflecting the stress-response state (which genes were activated, which pathways were triggered)
- Reverse transcriptase copies this stress-response RNA back into DNA and inserts it into the genome as a new genetic patch
- This is trial-and-error adaptation: Most insertions are neutral or harmful (cell dies), but occasionally one enhances stress-resistance (cell survives)
Step 5: The New Sequence Is Transcribed In Future Stress Events
- The inserted DNA becomes part of the genome (in non-coding regions)
- When the cell or its descendants encounter the same class of environmental stress, those sequences are transcribed again
- The RNA produced is a hybrid sequence—it reflects both:
- The cell's native stress-response machinery
- The specific toxin-breakdown signature from the original exposure
Why The "Viral RNA" Is Not Found In The Human Reference Genome:
Because it was just created during the current environmental-exposure event through reverse transcription and transposable-element activation.
The sequence is:
- ✓ Generated by human cellular machinery (reverse transcriptase, RNA polymerase, proteasome, MHC)
- ✓ A record of the specific toxin encountered (breakdown signature incorporated into stress-response RNA)
- ✓ Unique to this exposure event (novel combination reflecting this specific insult)
- ✗ NOT from the standard protein-coding genome (comes from non-coding "junk DNA" regions—98% of the genome—that are highly variable and responsive)
- ✗ NOT from an external organism (entirely endogenous process)
Analogy:
If you burn a new type of plastic in your house, the smoke will have a chemical signature that doesn't match any previous smoke in your records. But that doesn't mean a Smoke Organism invaded your house. It means combustion of that specific material produced a novel combination of chemicals.
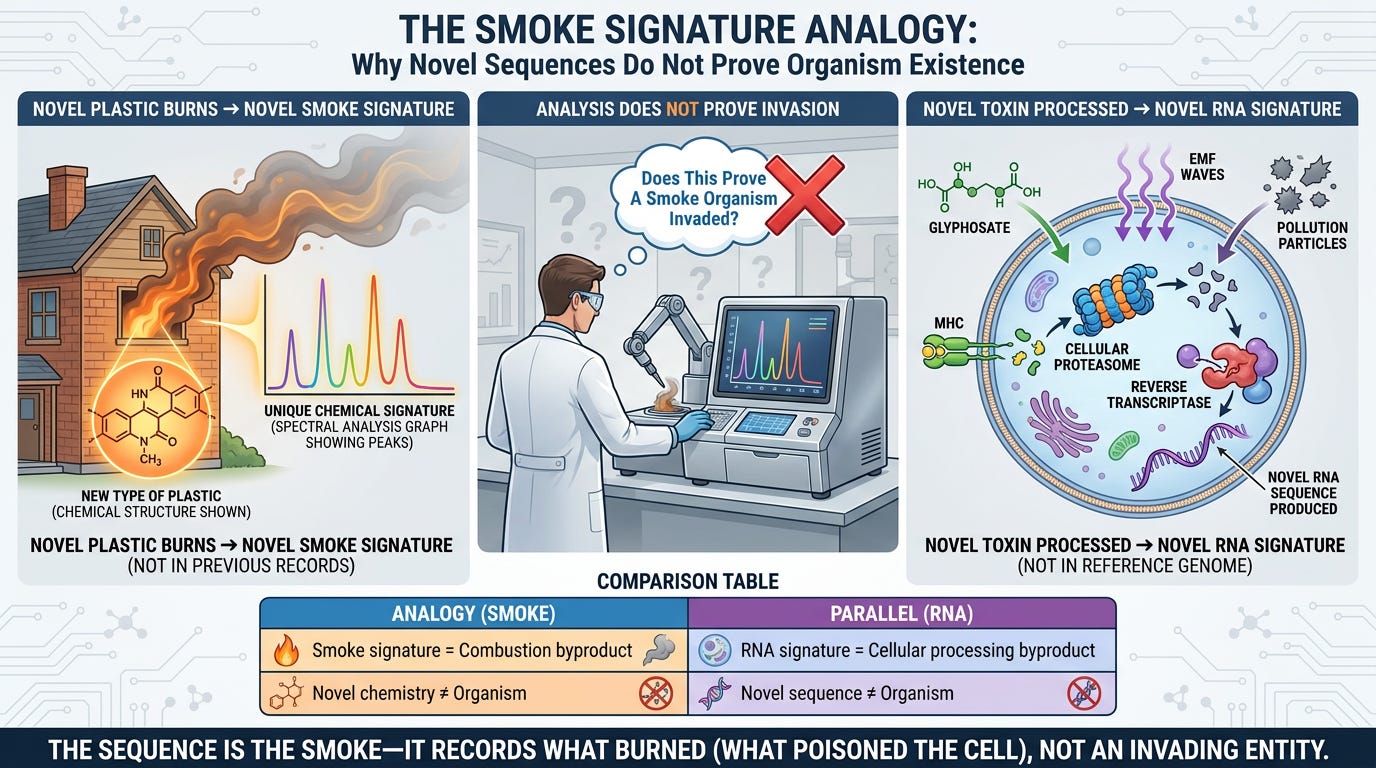
The RNA sequence is the smoke signature of the toxin being processed. It's novel because this specific toxin + this person's cellular processing = unique breakdown signature.
Why The Same Sequence Appears Across Different People (In Natural Exposure):
Because humans are humans. They have the same cellular machinery, the same stress-response pathways, and the same MHC processing systems.
When exposed to the same environmental insult, they produce convergent RNA signatures.
Scenario: 100 people in a city are exposed to the same insult (60GHz 5G rollout, glyphosate contamination, pollution spike).
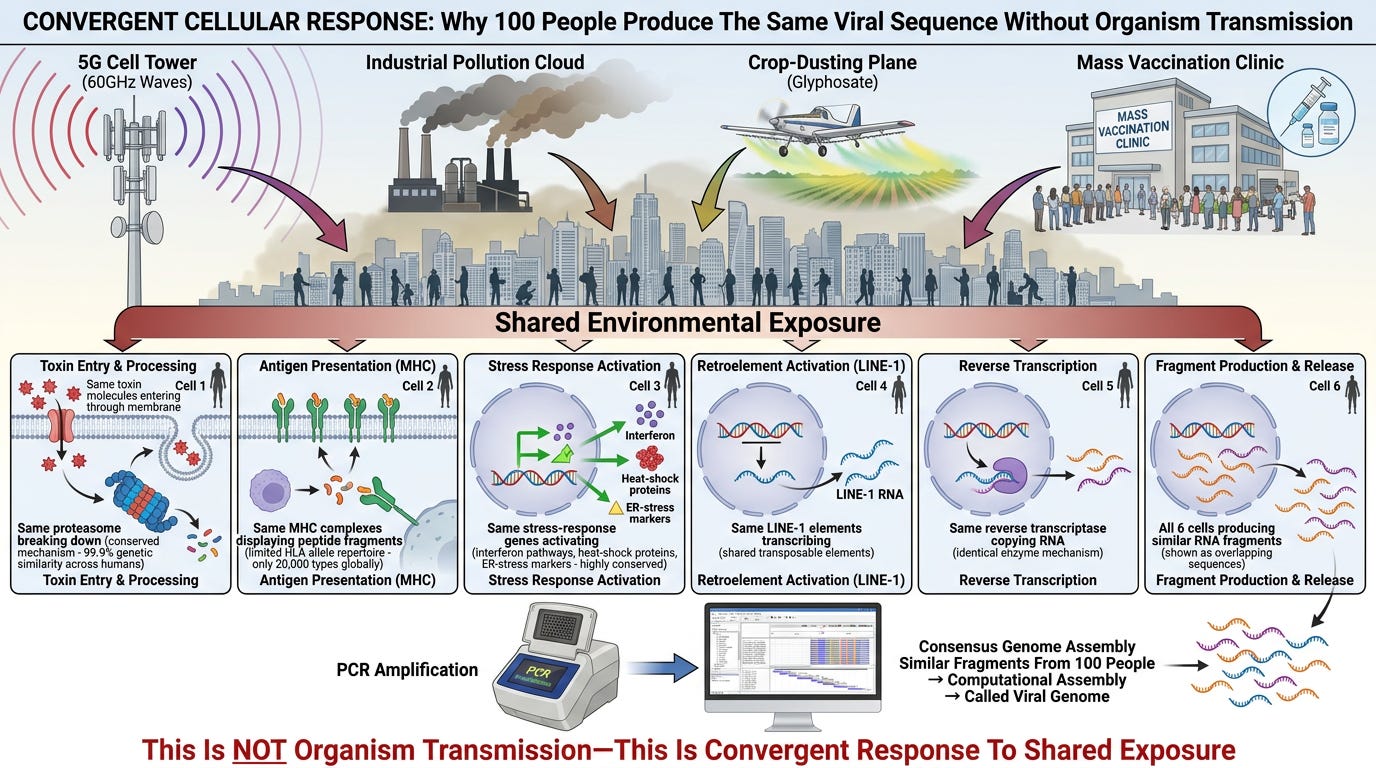
What happens in each person:
1. Same toxin enters cells (shared environmental exposure)
2. Same cellular processing pathways activate:
- Proteasome shreds using conserved enzymatic mechanisms (humans share 99.9% genetic similarity)
- MHC complexes (limited by HLA allele repertoire—only ~20,000 distinct HLA types exist across all humans)
- Stress-response genes (interferon pathways, heat-shock proteins, ER-stress markers)—highly conserved across humans
3. Same LINE-1 elements activate (transposable elements are shared across the population)
4. Reverse transcriptase copies similar stress-response RNAs
5. Convergent sequences emerge—not identical, but highly similar because:
- Same toxin → same breakdown fragments
- Same MHC processing → same peptide selection
- Same stress pathways → same gene activation
- Same reverse transcriptase → same copying mechanism
Result: When you sequence RNA from these 100 patients and assemble it computationally, you get a consensus sequence that appears "shared."
This is NOT organism transmission. This is convergent cellular response to shared environmental exposure.
SECTION SEVEN: Non-Coding DNA—The Adaptive Environmental-Response Library
The Old Paradigm: "Junk DNA"
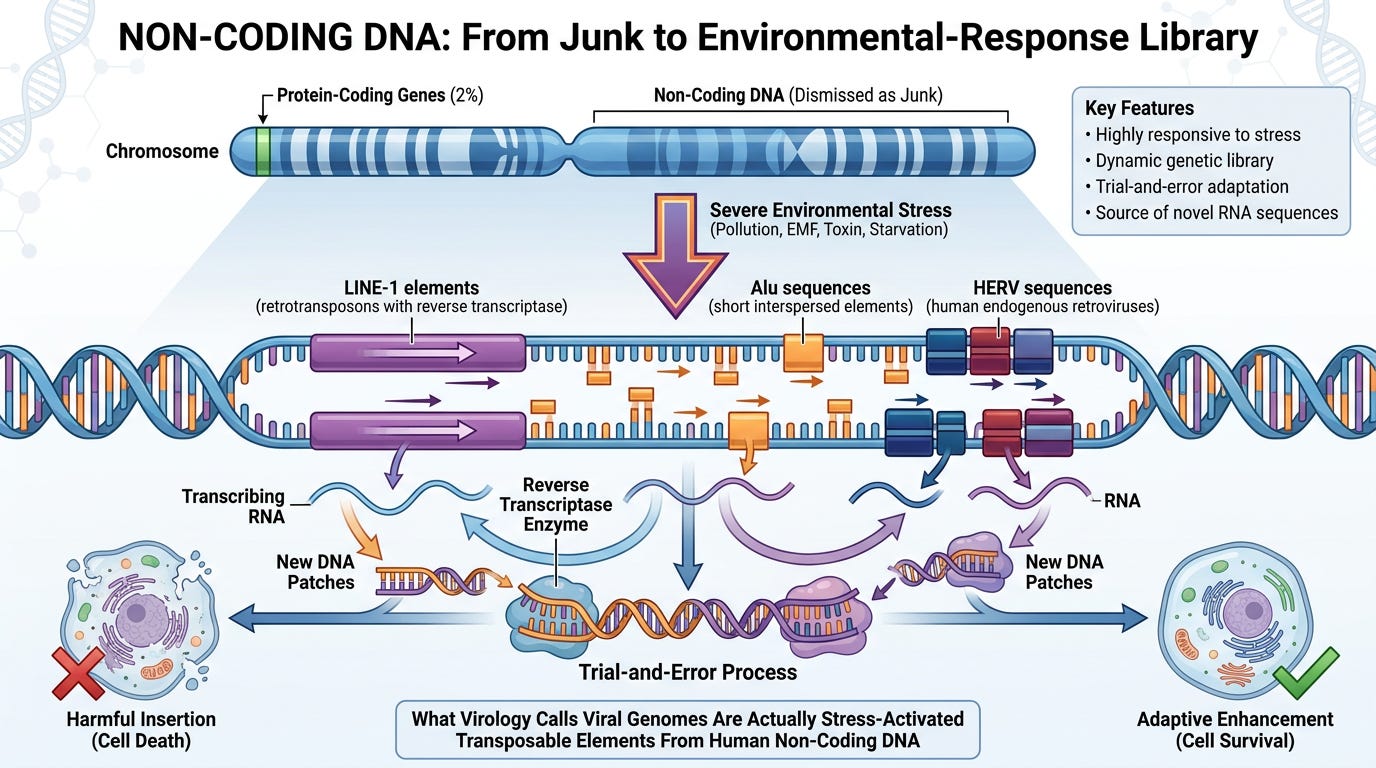
- 98% of the human genome does not code for proteins
- Early genetics dismissed this as evolutionary leftovers or "junk"
The New Understanding:
Non-coding DNA is:
- ✓ Highly responsive to environmental stress
- ✓ Contains transposable elements (LINE-1, Alu, HERV sequences—ancient viral-like sequences integrated millions of years ago)
- ✓ A dynamic genetic library for trial-and-error adaptation
- ✓ Activated during severe environmental exposure
- ✓ The source of "novel" RNA sequences virology calls "viral genomes" (in natural exposure scenarios)
Under severe environmental stress:
1. Severe insult (pollution, EMF, toxin, nutritional deficiency) hits population
2. Cells activate LINE-1 and Alu elements (transposable elements in non-coding DNA)
3. These elements transcribe themselves using reverse transcriptase
4. The RNA produced contains:
- Transposable-element sequences (LINE-1, Alu, HERV)
- Stress-response gene transcripts (interferons, heat-shock proteins)
- Toxin-breakdown signatures (from MHC processing)
5. PCR primers detect these sequences
6. Computational assembly creates a "genome" from these fragments
7. Virology labels it "novel viral genome"
8. Terrain framework recognizes it as: Endogenous transposable-element activation + toxin-response signature
This mechanism explains "novel" sequences in natural environmental exposure—but does NOT apply to reverse genetics, where you're simply detecting the synthetic construct you forcibly introduced.
SECTION EIGHT: Summary—What This Explains
Observation 1: "Viral" RNA detected in sick patients but not healthy controls
Virology: Virus present in sick, absent in healthy.
Terrain (Natural Exposure): Sick patients under environmental stress → transposable elements activated → stress-response RNA produced. Healthy controls not stressed → no activation.
Terrain (Lab Reverse Genetics): Sick cultures were force-fed synthetic construct. Healthy controls were not.
Observation 2: The "viral genome" contains sequences not in human protein-coding genes
Virology: Proves it's foreign, from an organism.
Terrain (Natural Exposure): Comes from non-coding transposable elements (98% of genome) activated during stress—human-derived but not protein-coding. Novel combinations from toxin-breakdown + reverse transcription.
Terrain (Lab Reverse Genetics): It's the lab-manufactured synthetic construct you introduced—of course it's not in the human genome, you chemically built it from the computationally-assembled fiction.
Observation 3: Same "viral sequence" appears across multiple patients in an outbreak
Virology: Organism transmitted person-to-person.
Terrain (Natural Exposure): Convergent cellular response—same environmental insult + same human cellular machinery = similar stress-response RNA across population.
Terrain (Lab Reverse Genetics): Not applicable—this is an artificial lab scenario, not a natural outbreak.
Observation 4: "Viral" RNA increases over time during "infection"
Virology: Virus replicates exponentially.
Terrain (Natural Exposure): Cellular stress escalates → more transposable-element activation → more stress-response RNA produced → more exosomes carrying these sequences.
Terrain (Lab Reverse Genetics): Cells are copying the synthetic input and packaging it into exosomes—cellular copying, not organism replication.
Observation 5: Reverse genetics using the "viral genome" sequence reproduces disease
Virology: Proves the genome is real and belongs to a virus.
Terrain: The synthetic sequence was reverse-engineered from cellular stress-response RNA (Scenario A). Reintroducing it (Scenario B) reproduces the cellular stress (UPR, exosome production, inflammation) in already-starved cells. You've re-introduced a toxic stimulus into a compromised system, which re-triggers the stress response. Perfect circularity. No organism validated.
Observation 6: No single particle ever shown to contain the full genome
Virology: Claims it's "technically impossible" to isolate.
Terrain: This is the admission that proves the organism was never independently demonstrated. In BOTH natural exposure and lab reverse genetics, the "genome" only exists as:
- A computational assembly (software output)
- A synthetic lab construct (chemical manufacturing output)
It never exists as a single intact molecule inside a single purified particle. This is the proof the organism claim is unfounded.
CONCLUSION: The Organism Assumption Was Never Validated
Reverse Genetics is the ultimate demonstration of virology's fundamental inversion: mistaking the cell's defensive response for an invading entity.
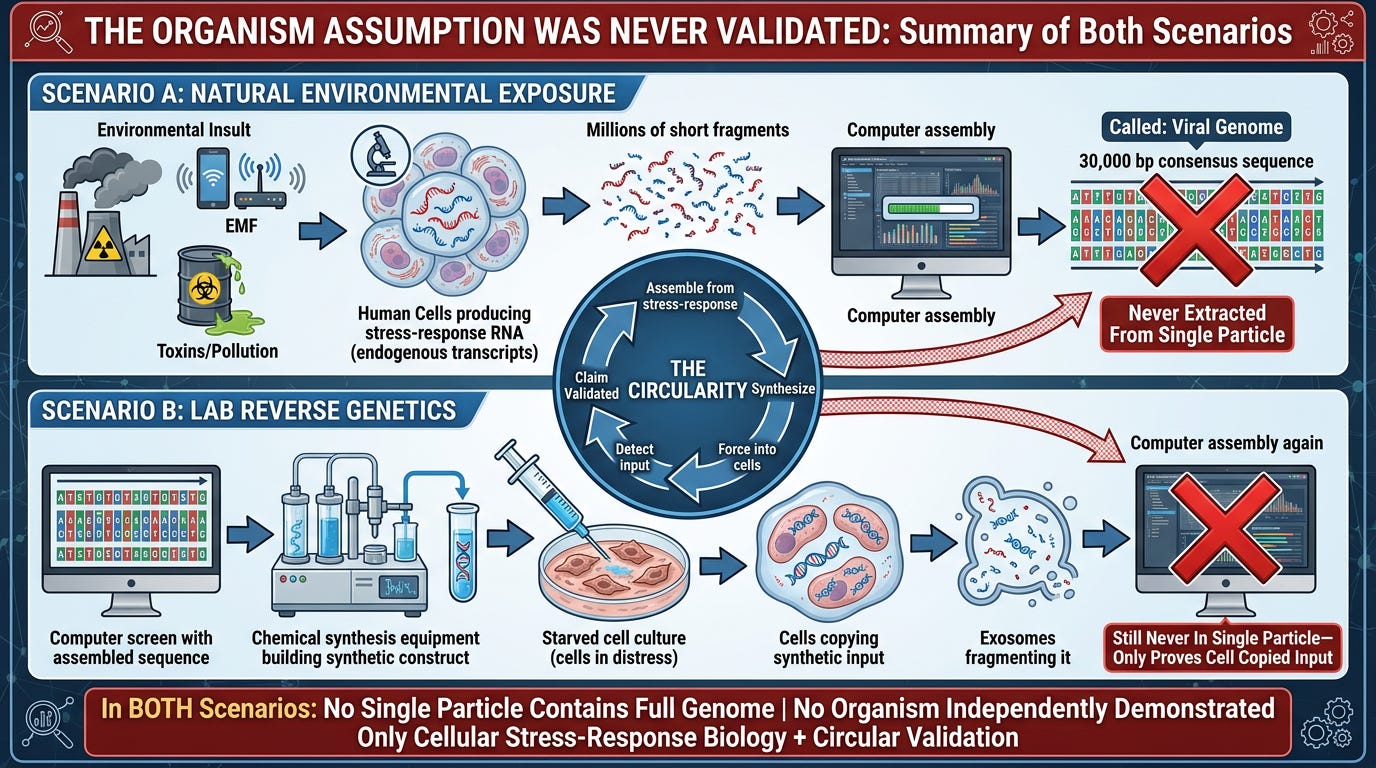
The data is clear, the biochemistry is precise, and the mechanism is well-documented:
In Natural Environmental Exposure (Scenario A):
1. Environmental insult occurs (pollution, EMF, toxin, vaccination, nutritional deficiency)
2. Cells produce stress-response RNA (endogenous transcripts from interferon genes, heat-shock proteins, transposable elements)
3. Toxin-breakdown signatures created through proteasome processing, MHC display, and reverse transcription
4. PCR detects these real sequences
5. Computational assembly stitches fragments into a "genome" that never existed as a single intact molecule
6. This is mislabeled "viral genome"
What it actually is: Environmental-insult signature—the cell's record of what poisoned it
In Laboratory Reverse Genetics (Scenario B):
1. Take the computationally-assembled fiction from Scenario A
2. Chemically synthesize the entire sequence base-by-base
3. Force it into starved, antibiotic-poisoned, isolated cell cultures
4. Cells transcribe, translate, and copy it (using their own machinery)
5. Cells fragment it for exosomal packaging and elimination
6. PCR detects the synthetic input (because primers were designed to match it)
7. Scientists claim "validation"
What it actually is: Cellular copying of a synthetic input + detection of what was forcibly introduced. Even here, no single particle contains the full genome.
The Perfect Circularity:
Step 1: Natural exposure → cellular stress-response → computational assembly → labeled "viral genome"
Step 2: Synthesize labeled genome → force into cells → cells copy it → detect it → "validated!"
This is not validation. This is circular reasoning masquerading as science.
The Unified Terrain Explanation:
In BOTH scenarios:
- No single particle ever contains the full genome
- Sequencing always produces millions of short reads
- Software always assembles the "genome"
- The organism assumption is never independently validated
What virology calls:
- "Viral entry" = Forced absorption of synthetic chemical (lab) OR environmental toxin exposure (natural)
- "Viral replication" = Cellular copying of input (lab) OR high-volume exosomal stress-response (natural)
- "Viral genome" = Synthetic construct you introduced (lab) OR computational assembly of stress-response transcripts (natural)
- "Viral shedding" = Cellular purging of toxic overload via exosomes (both)
- "Cytopathic effect" = Cellular death from starvation + toxin accumulation (lab) OR stress-response in living organism (natural)
The Terrain Framework accounts for all observations in both scenarios. The organism claim remains unproven in both.
The burden of proof has never been met.
Cross-References:
§§ 1.2, 1.15, 5.1, 5.6, 5.7.A-C (The Open Reality Bible)
"The Gain-of-Function Hijack: How Molecular Biology Got Relabeled as Virus Enhancement" (May 22, 2026)
"The Virus Audit: A Complete Physical-Evidence Falsification of Modern Virology" (May 18, 2026)
This article is part of a comprehensive physical-evidence audit of modern virology. All claims are grounded in peer-reviewed molecular biology, cellular biochemistry, and published experimental protocols. The Terrain Framework offers a biochemically complete explanation that requires no unproven entity assumptions.
I have added a link to this article to a few people's posts in the hope that they will seek you out, Shan. xx